
Hjernebarken - Cerebral cortex
| Hjernebarken | |
|---|---|
 Vevskive fra hjernen til en voksen makakape ( Macaca mulatta ). Hjernebarken er det ytre laget avbildet i mørkt fiolett. Kilde: BrainMaps.org
| |

Golgi-fargede nevroner i cortex
| |
| Detaljer | |
| Del av | Cerebrum |
| Identifikatorer | |
| Latin | Cortex cerebri |
| MeSH | D002540 |
| NeuroNames | 39 |
| NeuroLex ID | birnlex_1494 |
| TA98 |
A14.1.09.003 A14.1.09.301 |
| TA2 | 5527 , 5528 |
| FMA | 61830 |
| Anatomiske termer for neuroanatomi | |
Den cerebrale cortex , også kjent som cerebral mantelen , er det ytre laget av nervevev i cerebrum av hjernen i mennesker og andre pattedyr . Hjernebarken består for det meste av seks-lags neocortex , med bare 10% bestående av allocortex . Den er delt inn i to cortices , av den langsgående sprekken som deler cerebrum i venstre og høyre cerebral halvkule . De to halvkule er forbundet under cortex av corpus callosum . Hjernebarken er det største stedet for nevral integrasjon i sentralnervesystemet . Det spiller en nøkkelrolle i oppmerksomhet , persepsjon , bevissthet , tanke , minne , språk og bevissthet .
I de fleste pattedyr, bortsett fra små pattedyr som har små hjerner, er hjernebarken foldet, noe som gir et større overflateareal i det avgrensede volum av kraniet . Bortsett fra å minimere hjerne- og kranialvolumet, er kortikalfolding avgjørende for ledningen til hjernen og dens funksjonelle organisering. Hos pattedyr med en liten hjerne er det ingen folding og cortex er glatt.
En fold eller ås i cortex kalles en gyrus (flertall gyri) og et spor kalles en sulcus (flertall sulci). Disse overflatekonvolusjonene vises under fosterutvikling og fortsetter å modnes etter fødselen gjennom prosessen med gyrifikasjon . I menneskehjernen er størstedelen av hjernebarken ikke synlig utenfra, men begravet i sulci, og den øsformede cortex er helt skjult. De store sulci og gyri markerer delingene av lillehjernen i hjernelappene .
Det er mellom 14 og 16 milliarder nevroner i den menneskelige hjernebarken. Disse er organisert i horisontale lag, og radielt i kortikale kolonner og minikolonner . Kortikale områder har spesifikke funksjoner som bevegelse i motorisk cortex og syn i visuell cortex .
Struktur

Hjernebarken er det ytre dekket av overflatene på hjernehalvdelene og brettes til topper som kalles gyri , og spor som kalles sulci . I menneskehjernen er den mellom to og tre eller fire millimeter tykk, og utgjør 40 prosent av hjernens masse. 90 prosent av hjernebarken er seks-lags neocortex med de andre 10 prosentene som består av allocortex . Det er mellom 14 og 16 milliarder nevroner i cortex, og disse er organisert radielt i kortikale kolonner og minikolonner i de horisontalt organiserte lagene i cortex.
Neocortex er separerbar i forskjellige områder av cortex kjent i flertall som cortices, og inkluderer motor cortex og visual cortex . Omtrent to tredjedeler av den kortikale overflaten er begravet i sulci og den insulære cortex er helt skjult. Cortex er tykkest over toppen av en gyrus og tynnest i bunnen av en sulcus.
Bretter
Hjernebarken er foldet på en måte som gjør det mulig for et stort overflateareal på neuralt vev for å passe innenfor rammen av neurocranium . Når den brettes ut i mennesket, har hver hemisfærisk cortex et totalt overflateareal på omtrent 0,12 kvadratmeter (1,3 kvadratmeter). Foldingen er innover vekk fra hjernens overflate, og er også tilstede på medialoverflaten på hver halvkule i den langsgående sprekken . De fleste pattedyr har en hjernebark som er innviklet med toppene kjent som gyri og trauene eller sporene kjent som sulci. Noen små pattedyr, inkludert noen små gnagere, har glatte hjerneoverflater uten gyrifisering .
Lober
De større sulci og gyri markerer divisjonene av hjernebarken i hjernen i lober i hjernen . Det er fire hovedlapper: frontallappen , parietallappen , tinninglappen og occipitallappen . Den insulære cortex er ofte inkludert som den insular lobe. Den limbiske lap er en kant av cortex på medialsiden av hver halvkule og er også ofte inkludert. Det er også beskrevet tre lobuler i hjernen: paracentral lobule , superior parietal lobule og inferior parietal lobule .
Tykkelse
For pattedyrarter har større hjerner (i absolutte termer, ikke bare i forhold til kroppsstørrelse) en tykkere cortice. De minste pattedyrene, som spissmus , har en neokortikal tykkelse på omtrent 0,5 mm; de med de største hjernene, som mennesker og finhvaler, har en tykkelse på 2–4 mm. Det er et omtrent logaritmisk forhold mellom hjernens vekt og kortikal tykkelse. Magnetisk resonansavbildning av hjernen (MRI) gjør det mulig å få et mål for tykkelsen på den menneskelige hjernebarken og relatere den til andre tiltak. Tykkelsen på forskjellige kortikale områder varierer, men generelt er sensorisk cortex tynnere enn motor cortex. En studie har funnet en positiv sammenheng mellom kortikal tykkelse og intelligens . En annen studie har funnet ut at den somatosensoriske cortex er tykkere hos migrene , selv om det ikke er kjent om dette er et resultat av migreneanfall eller årsaken til dem. En senere studie med en større pasientpopulasjon rapporterer ingen endring i kortikaltykkelsen hos migrene. En genetisk lidelse i hjernebarken, der redusert bretting i visse områder resulterer i en mikrogyrus , der det er fire lag i stedet for seks, er i noen tilfeller sett på å være relatert til dysleksi .
Lag av neocortex



Den neocortex er dannet av seks lag, nummerert fra I til VI, fra det ytterste lag I - i nærheten av den pia mater , til det innerste laget VI - i nærheten av den underliggende hvite substans . Hvert kortikale lag har en karakteristisk fordeling av forskjellige nevroner og deres forbindelser med andre kortikale og subkortikale regioner. Det er direkte forbindelser mellom forskjellige kortikale områder og indirekte forbindelser via thalamus.
Et av de tydeligste eksemplene på kortikal lagdeling er linjen til Gennari i den primære visuelle cortex . Dette er et bånd av hvitere vev som kan observeres med det blotte øye i calcarine sulcus i occipital lobe. Linjen til Gennari består av aksoner som bringer visuell informasjon fra thalamus til lag IV i den visuelle cortex .
Farging av tverrsnitt av cortex for å avsløre posisjonen til neuronale cellelegemer og de intrakortiske axon-traktene tillot nevroanatomene i begynnelsen av 1900-tallet å produsere en detaljert beskrivelse av cortexens laminære struktur hos forskjellige arter. Arbeidet til Korbinian Brodmann (1909) slo fast at neocortexen hos pattedyr konsekvent er delt inn i seks lag.
Lag I
Lag I er det molekylære laget , og inneholder få spredte nevroner, inkludert GABAergic rosehip neurons . Lag I består i stor grad av utvidelser av apikale dendritiske tufter av pyramidale nevroner og horisontalt orienterte aksoner, samt glialceller . Under utviklingen er Cajal-Retzius-celler og subpial granulære lagceller tilstede i dette laget. Noen spiny stellatceller kan også bli funnet her. Input til de apikale tuftene antas å være avgjørende for tilbakemeldingsinteraksjonene i hjernebarken som er involvert i assosiativ læring og oppmerksomhet. Selv om det en gang ble antatt at innspillet til laget jeg kom fra selve cortex, er det nå innsett at lag I over hjernebarkmantelen mottar betydelig input fra matrise- eller M-type thalamusceller (i motsetning til kjerne eller C-type som gå til lag IV).
Lag II
Lag II, det ytre granulære laget , inneholder små pyramideformede nevroner og mange stellate nevroner.
Lag III
Lag III, det ytre pyramidale laget , inneholder hovedsakelig små og mellomstore pyramidale nevroner, så vel som ikke-pyramidale nevroner med vertikalt orienterte intrakortikale aksoner; lag I til og med III er hovedmålet for interhemisfæriske kortikokortiske afferenter , og lag III er hovedkilden til kortikokortiske efferenter .
Lag IV
Lag IV, det indre granulære laget , inneholder forskjellige typer stellat- og pyramidale celler, og er hovedmålet for thalamokortiske afferenter fra thalamus type C-nevroner (kjernetype) samt intrahemisfæriske kortikokortiske afferenter. Lagene over lag IV omtales også som supragranulære lag (lag I-III), mens lagene nedenfor omtales som infragranulære lag (lag V og VI).
Lag V
Lag V, det indre pyramidale laget , inneholder store pyramidale nevroner. Aksoner fra disse forlater cortex og får forbindelse med subkortikale strukturer inkludert basale ganglier . I den primære motoriske cortex i frontallappen inneholder lag V gigantiske pyramideceller kalt Betz -celler , hvis aksoner beveger seg gjennom den indre kapsel , hjernestammen og ryggmargen som danner kortikospinalkanalen , som er hovedveien for frivillig motorisk kontroll .
Lag VI
Lag VI, det polymorfe eller multiforme laget, inneholder få store pyramidale nevroner og mange små spindellignende pyramidale og multiforme nevroner; lag VI sender efferente fibre til thalamus, og etablerer en veldig presis gjensidig sammenkobling mellom cortex og thalamus. Det vil si at lag VI -nevroner fra en kortikal kolonne kobles til thalamus -nevroner som gir innspill til den samme kortikale kolonnen. Disse forbindelsene er både eksitatoriske og hemmende. Nevroner sender eksitatoriske fibre til nevroner i thalamus og sender også kollateraler til thalamisk retikulær kjerne som hemmer de samme thalamus -nevronene eller de som grenser til dem. En teori er at fordi den hemmende produksjonen reduseres av kolinerge innganger til hjernebarken, gir dette hjernestammen justerbar "forsterkningskontroll for reléet til lemniscale innganger".
Kolonner
De kortikale lagene er ikke bare stablet over hverandre; det eksisterer karakteristiske forbindelser mellom forskjellige lag og nevronale typer, som strekker seg over hele tykkelsen av cortex. Disse kortikale mikrokretsene er gruppert i kortikale kolonner og minikolonner . Det har blitt foreslått at minikolonnene er de grunnleggende funksjonelle enhetene i cortex. I 1957 viste Vernon Mountcastle at de funksjonelle egenskapene til cortex endrer seg brått mellom lateralt tilstøtende punkter; de er imidlertid kontinuerlige i retningen vinkelrett på overflaten. Senere arbeider har gitt bevis på tilstedeværelsen av funksjonelt distinkte kortikale kolonner i den visuelle cortex (Hubel og Wiesel , 1959), auditiv cortex og associative cortex.
Kortikale områder som mangler et lag IV kalles agranular . Kortikale områder som bare har et rudimentært lag IV kalles dysgranular. Informasjonsbehandling innenfor hvert lag er bestemt ved ulike tids- dynamikk med at det i sjiktene II / III med en langsom 2 Hz oscillasjon mens det i sjiktet V har en rask 10-15 Hz oscillasjon.
Typer av cortex
Basert på forskjellene i laminær organisering kan hjernebarken klassifiseres i to typer, det store området med neocortex som har seks cellelag, og det mye mindre området av allocortex som har tre eller fire lag:
- Neocortex er også kjent som isocortex eller neopallium og er den delen av den modne hjernebarken med seks forskjellige lag. Eksempler på neokortikale områder inkluderer den granulære primære motoriske cortex , og den striate primære visuelle cortex . Neocortex har to undertyper, den sanne isocortex og proisocortex som er en overgangsregion mellom isocortex og regionene i periallocortex.
- Allocortex er den delen av hjernebarken med tre eller fire lag, og har tre undertyper, paleocortex med tre kortikale laminae, archicortex som har fire eller fem, og et overgangsområde ved siden av allocortex, periallocortex . Eksempler på allocortex er luktbarken og hippocampus .
Det er et overgangsområde mellom neocortex og allocortex kalt paralimbic cortex , hvor lag 2, 3 og 4 slås sammen. Dette området inneholder proisocortex av neocortex og periallocortex av allocortex. I tillegg kan hjernebarken klassifiseres i fire lapper : frontallappen , tinninglappen , parietallappen og occipitallappen , navngitt fra deres overliggende bein i skallen.
Blodtilførsel og drenering

Blodtilførsel til hjernebarken er en del av hjernens sirkulasjon . Cerebrale arterier forsyner blodet som perfuserer cerebrum. Dette arterielle blodet fører oksygen, glukose og andre næringsstoffer til cortex. Cerebrale vener drenerer det deoksygenerte blodet, og metabolsk avfall inkludert karbondioksid, tilbake til hjertet.
De viktigste arteriene som forsyner cortex er den fremre cerebrale arterien , den midtre cerebrale arterien og den bakre cerebrale arterien . Den fremre cerebrale arterien forsyner de fremre delene av hjernen, inkludert det meste av frontallappen. Den midtre cerebrale arterie forsyner parietallappene, temporallappene og deler av occipital -lappene. Den midtre cerebrale arterien deler seg i to grener for å forsyne venstre og høyre halvkule, hvor de forgrener seg videre. Den bakre cerebrale arterien forsyner occipital lappene.
Den sirkelen av Willis er hovedblodsystemet som omhandler blodtilførselen i storhjernen og hjernebarken.

Utvikling
Den prenatale utviklingen av hjernebarken er en kompleks og finjustert prosess kalt kortikogenese , påvirket av samspillet mellom gener og miljø.
Nevralrør
Hjernebarken utvikler seg fra den mest fremre delen, forhjerneområdet, i nevralrøret . De neurale plate folder og lukkes for å danne det nevrale røret . Fra hulrommet inne i nevralrøret utvikler ventrikkelsystemet , og fra neuroepitelcellene i veggene neuronene og glia i nervesystemet. Den mest fremre (forreste eller kraniale) delen av nevralplaten , prosencephalon , som er tydelig før nevralisering begynner, gir opphav til hjernehalvdelene og senere cortex.
Kortikal nevronutvikling
Kortikale nevroner genereres i ventrikkelsonen , ved siden av ventriklene . Først inneholder denne sonen nevrale stamceller , som overgår til radiale glialceller - stamceller, som deler seg for å produsere glialceller og nevroner.
Radial glia

Hjernebarken består av en heterogen populasjon av celler som gir opphav til forskjellige celletyper. Flertallet av disse cellene er avledet fra radial glia -migrasjon som danner de forskjellige celletypene i neocortex, og det er en periode forbundet med en økning i nevrogenese . På samme måte regulerer prosessen med nevrogenese laminering for å danne de forskjellige lagene i cortex. I løpet av denne prosessen er det en økning i begrensningen av celleskjebne som begynner med at tidligere forfedre gir opphav til hvilken som helst celletype i cortex og senere forfedre som bare gir opphav til nevroner av overfladiske lag. Denne differensielle celleskjebnen skaper en innvendig og utvendig topografi i cortex med yngre nevroner i overfladiske lag og eldre nevroner i dypere lag. I tillegg stoppes laminære nevroner i S- eller G2 -fasen for å gi et godt skille mellom de forskjellige kortikale lagene. Laminar differensiering er ikke fullstendig fullført før etter fødselen siden laminære nevroner under utvikling fortsatt er følsomme for ytre signaler og miljømessige signaler.
Selv om flertallet av cellene som komponerer cortex stammer lokalt fra radial glia, er det en delmengde av nevroner som migrerer fra andre regioner. Radial glia gir opphav til nevroner som er pyramideformet og bruker glutamat som en nevrotransmitter , men disse migrerende cellene bidrar med nevroner som er stjerneformede og bruker GABA som deres viktigste nevrotransmitter. Disse GABAergiske nevronene genereres av stamceller i den mediale ganglioniske eminensen (MGE) som migrerer tangentielt til cortex via subventrikulær sone . Denne migrasjonen av GABAergiske nevroner er spesielt viktig siden GABA -reseptorer er eksitatoriske under utvikling. Denne eksitasjonen er først og fremst drevet av strømmen av kloridioner gjennom GABA -reseptoren, men hos voksne skifter kloridkonsentrasjoner som forårsaker en innadstrømning av klorid som hyperpolariserer postsynaptiske nevroner . Glialfibrene som produseres i de første divisjonene av stamcellene er radialt orientert, og strekker seg over tykkelsen av cortex fra ventrikkelsonen til den ytre pialoverflaten , og gir stillas for migrering av nevroner utover fra ventrikkelsonen .
Ved fødselen er det svært få dendritter tilstede på cortikalneurons cellekropp, og axonet er uutviklet. I løpet av det første leveåret blir dendritene dramatisk flere, slik at de kan romme opptil hundre tusen synaptiske forbindelser med andre nevroner. Axonet kan utvikle seg til å strekke seg langt fra cellelegemet.
Asymmetrisk inndeling
De første delene av stamcellene er symmetriske, som dupliserer det totale antallet stamceller ved hver mitotiske syklus . Deretter begynner noen stamceller å dele seg asymmetrisk og produserer en postmitotisk celle som vandrer langs de radiale glialfibrene, forlater ventrikkelsonen og en stamcelle, som fortsetter å dele seg til slutten av utviklingen, når den differensierer til en glialcelle eller en ependymal celle . Som G1 fase av mitose er langstrakt, i det som blir sett på som selektive cellesyklus-forlengelse, den nyfødte nevroner migrere til mer overfladiske lag av hjernebarken. De migrerende dattercellene blir pyramidale celler i hjernebarken. Utviklingsprosessen er tidsbestilt og regulert av hundrevis av gener og epigenetiske reguleringsmekanismer .
Lagorganisasjon

Den lagdelte strukturen til den modne hjernebarken dannes under utvikling. De første pyramidale nevronene som genereres migrerer ut av ventrikkelsonen og subventrikulær sone , sammen med reelinproduserende Cajal -Retzius -nevroner , fra forplaten . Deretter deler en kohorte av nevroner som migrerer inn i midten av forplaten dette forbigående laget i den overfladiske randsonen , som vil bli lag I i den modne neocortexen, og underplaten , som danner et mellomlag som kalles den kortikale platen . Disse cellene vil danne de dype lagene i den modne cortex, lag fem og seks. Senere fødte nevroner vandrer radielt inn i den kortikale platen forbi de dype lagneuronene og blir de øvre lagene (to til fire). Dermed blir lagene i cortex opprettet i en innvendig rekkefølge. Det eneste unntaket fra denne innvendige sekvensen av nevrogenese forekommer i laget I av primater , der neurogenese , i motsetning til gnagere , fortsetter gjennom hele perioden med kortikogenese .
Kortikal mønster

Kartet over funksjonelle kortikale områder, som inkluderer primær motorisk og visuell cortex, stammer fra en ' protomap ', som er regulert av molekylære signaler som fibroblastvekstfaktor FGF8 tidlig i embryonal utvikling. Disse signalene regulerer størrelsen, formen og plasseringen av kortikale områder på overflaten av det kortikale primordium, delvis ved å regulere gradienter av transkripsjonsfaktoruttrykk , gjennom en prosess som kalles kortikal mønster . Eksempler på slike transkripsjonsfaktorer inkluderer genene EMX2 og PAX6 . Til sammen danner begge transkripsjonsfaktorene en motsatt uttrykksgradient. Pax6 er sterkt uttrykt ved den rostrale laterale polen, mens Emx2 er sterkt uttrykt i den kaudomediale polen. Etableringen av denne gradienten er viktig for riktig utvikling. For eksempel kan mutasjoner i Pax6 få ekspresjonsnivåer av Emx2 til å ekspandere ut av det normale ekspresjonsdomenet, noe som til slutt ville føre til en utvidelse av områdene som normalt stammer fra den caudale mediale cortex, for eksempel den visuelle cortex . Tvert imot, hvis mutasjoner i Emx2 oppstår, kan det føre til at Pax6-uttrykkende domene utvides og resultere i at frontale og motoriske kortikale regioner forstørres. Derfor tror forskere at lignende gradienter og signalsentre ved siden av cortex kan bidra til det regionale uttrykket for disse transkripsjonsfaktorene. To meget godt studerte mønstersignaler for cortex inkluderer FGF og retinsyre . Hvis FGF -er misuttrykkes i forskjellige områder av utviklingsbarken, blir kortikal mønster forstyrret. Spesielt når Fgf8 økes i den fremre polen, blir Emx2 nedregulert og et kaudalt skifte i den kortikale regionen oppstår. Dette fører til syvende og sist til en utvidelse av rostralregionene. Derfor spiller Fgf8 og andre FGFer en rolle i reguleringen av uttrykk for Emx2 og Pax6 og representerer hvordan hjernebarken kan bli spesialisert for forskjellige funksjoner.
Rask utvidelse av det kortikale overflatearealet reguleres av mengden selvfornyelse av radiale glialceller og er delvis regulert av FGF- og Notch-gener . I perioden med kortikal nevrogenese og lagdannelse begynner mange høyere pattedyr prosessen med gyrifikasjon , som genererer de karakteristiske brettene i hjernebarken. Gyrifikasjon reguleres av et DNA-assosiert protein Trnp1 og av FGF- og SHH- signalering
Utvikling
Av alle de forskjellige hjerneområdene viser hjernebarken den største evolusjonære variasjonen og har utviklet seg sist. I motsetning til den svært bevarte kretsen til for eksempel medulla oblongata , som tjener kritiske funksjoner som regulering av hjerte og respirasjonsfrekvenser, er mange områder av hjernebarken ikke strengt nødvendige for å overleve. Dermed har utviklingen av hjernebarken oppdaget og modifisert nye funksjonelle områder - spesielt assosiasjonsområder som ikke direkte mottar innspill fra utenfor cortex.
En nøkkel teori av kortikal evolusjon er nedfelt i den hypotese om radiale enheter og tilhørende protomap hypotese, først foreslått av Rakic. Denne teorien sier at nye kortikale områder dannes ved tillegg av nye radiale enheter, som oppnås på stamcellenivå . Protomap -hypotesen sier at den cellulære og molekylære identiteten og egenskapene til nevroner i hvert kortikale område er spesifisert av kortikale stamceller , kjent som radiale glialceller , i et urkart. Dette kartet blir styrt av utskilte signaliseringsproteiner og nedstrøms transkripsjonsfaktorer .
Funksjon

Tilkoblinger
Hjernebarken er koblet til forskjellige subkortikale strukturer som thalamus og basalganglier , sender informasjon til dem langs efferente forbindelser og mottar informasjon fra dem via afferente forbindelser . Mest sanseinformasjon blir dirigert til hjernebarken via thalamus. Olfaktorisk informasjon passerer imidlertid gjennom luktpæren til olfaktorisk cortex ( piriform cortex ). Flertallet av forbindelsene er fra ett område av cortex til et annet, i stedet for fra subkortiske områder; Braitenberg og Schüz (1998) hevder at i primære sanseområder, på det kortikale nivået hvor inputfibrene avsluttes, blir opptil 20% av synapser levert av ekstrakortikale afferenter, men at i andre områder og andre lag sannsynligvis vil andelen være mye Nedre.
Kortikale områder
Hele hjernebarken ble delt inn i 52 forskjellige områder i en tidlig presentasjon av Korbinian Brodmann . Disse områdene kjent som Brodmann -områder , er basert på deres cytoarchitecture, men er også knyttet til forskjellige funksjoner. Et eksempel er Brodmann -område 17 som er den primære visuelle cortex .
I mer generelle termer beskrives cortex vanligvis som å omfatte tre deler: sensoriske, motoriske og assosieringsområder.
Sensoriske områder


De sensoriske områdene er de kortikale områdene som mottar og behandler informasjon fra sansene . Deler av cortex som mottar sensoriske innganger fra thalamus kalles primære sensoriske områder. Sansene for syn, hørsel og berøring betjenes av henholdsvis primær visuell cortex, primær auditiv cortex og primær somatosensorisk cortex . Generelt mottar de to halvkule informasjon fra den motsatte (kontralaterale) siden av kroppen . For eksempel mottar høyre primære somatosensoriske cortex informasjon fra venstre lemmer, og høyre visuelle cortex mottar informasjon fra venstre synsfelt . Organiseringen av sensoriske kart i cortex gjenspeiler den til det tilsvarende sanseorganet, i det som er kjent som et topografisk kart . Nabopunkter i den primære visuelle cortex , for eksempel, tilsvarer nabopunkter i netthinnen . Dette topografiske kartet kalles et retinotopisk kart . På samme måte eksisterer det et tonotopisk kart i den primære auditive cortex og et somatotopisk kart i den primære sensoriske cortex. Dette siste topografiske kartet over kroppen til den sentrale gyrusen bak har blitt illustrert som en deformert menneskelig representasjon, den somatosensoriske homunculus , hvor størrelsen på forskjellige kroppsdeler gjenspeiler den relative tettheten av deres innervering. Områder med mye sensorisk innervasjon, for eksempel fingertuppene og leppene, krever mer kortikale områder for å behandle finere sansning.
Motorområder
Motorområdene er plassert i begge halvkule av cortex. Motorområdene er veldig nært knyttet til kontroll av frivillige bevegelser, spesielt fine fragmenterte bevegelser utført av hånden. Den høyre halvdelen av motorområdet styrer venstre side av kroppen, og omvendt.
To områder av cortex kalles vanligvis motor:
- Primær motorisk cortex , som utfører frivillige bevegelser
- Supplerende motoriske områder og premotorisk cortex , som velger frivillige bevegelser.
I tillegg er motorfunksjoner beskrevet for:
- Posterior parietal cortex , som leder frivillige bevegelser i rommet
- Dorsolateral prefrontal cortex , som bestemmer hvilke frivillige bevegelser som skal utføres i henhold til instruksjoner, regler og selvgenererte tanker av høyere orden.
Like under hjernebarken er sammenkoblede subkortikale masser av grå materie kalt basale ganglier (eller kjerner). Basalganglier mottar innspill fra substantia nigra i mellomhjernen og motorområdene i hjernebarken, og sender signaler tilbake til begge disse stedene. De er involvert i motorstyring. De finnes lateralt til thalamus. Hovedkomponentene i basalgangliene er caudatkjernen , putamen , globus pallidus , substantia nigra , nucleus accumbens og subthalamic nucleus . Putamen og globus pallidus er også samlet kjent som lentiformkjernen , fordi de sammen danner en linseformet kropp. Putamen og caudate -kjernen kalles også kollektivt corpus striatum etter deres stripete utseende.
Foreningsområder

Foreningsområdene er de delene av hjernebarken som ikke tilhører de primære regionene. De fungerer for å produsere en meningsfull opplevelse av verden, sette oss i stand til å samhandle effektivt og støtte abstrakt tenkning og språk. Den parietal , temporal , og occipital lobes - alt ligger i den bakre delen av hjernebarken - integrere sanseinformasjon og informasjon som er lagret i minnet. Den frontallappen eller prefrontal foreningen komplekset er involvert i planlegging av tiltak og bevegelse, samt abstrakt tenkning. Globalt sett er foreningsområdene organisert som distribuerte nettverk. Hvert nettverk forbinder områder fordelt på områder med store mellomrom i cortex. Distinkte nettverk er plassert ved siden av hverandre og gir en kompleks serie med sammenvevde nettverk. Den spesifikke organisasjonen av foreningsnettverkene debatteres med bevis for interaksjoner, hierarkiske forhold og konkurranse mellom nettverk.
Hos mennesker er foreningsnettverk spesielt viktige for språkfunksjonen. Tidligere ble det teoretisert at språklige evner er lokalisert i Brocas område i områder av venstre inferior frontal gyrus , BA44 og BA45 , for språkuttrykk og i Wernickes område BA22 , for språkmottak . Imidlertid har prosessene for språkuttrykk og mottak vist seg å forekomme i andre områder enn bare de strukturene rundt lateral sulcus , inkludert frontallappen, basalganglia , lillehjernen og pons .
Klinisk signifikans

Neurodegenerative sykdommer som Alzheimers sykdom og Lafora sykdom , viser som en markør en atrofi av den grå substansen i hjernebarken.
Andre sykdommer i sentralnervesystemet inkluderer nevrologiske lidelser som epilepsi , bevegelsesforstyrrelser og talevansker ( afasi ).
Hjerneskade fra sykdom eller traumer, kan innebære skade på en spesifikk lapp, for eksempel i frontallapplidelse , og tilhørende funksjoner vil bli påvirket. Den blod-hjernebarrieren som tjener til å beskytte hjerne fra infeksjon kan bli kompromittert og tillate innføring til patogener .
Den fosteret er utsatt for en rekke miljøfaktorer som kan føre til fødselsskader og problemer i senere utvikling. Mors alkoholforbruk kan for eksempel forårsake fostrets alkoholspekterforstyrrelse . Andre faktorer som kan forårsake nevroutviklingsforstyrrelser er toksiske stoffer som medisiner og eksponering for stråling fra røntgenstråler . Infeksjoner kan også påvirke utviklingen av cortex. En virusinfeksjon er en av årsakene til lissencephaly , noe som resulterer i en jevn cortex uten gyrifikasjon .
En type elektrokortikografi som kalles kortikal stimuleringskartlegging er en invasiv prosedyre som innebærer å plassere elektroder direkte på den eksponerte hjernen for å lokalisere funksjonene til bestemte områder av cortex. Det brukes i kliniske og terapeutiske applikasjoner, inkludert prekirurgisk kartlegging.
Gener assosiert med kortikale lidelser
Det er en rekke genetiske mutasjoner som kan forårsake et bredt spekter av genetiske lidelser i hjernebarken, inkludert mikrocefali , schizencephaly og typer lissencephaly . Kromosomavvik kan også forårsake en rekke nevroutviklingsforstyrrelser som skjørt X -syndrom og Rett -syndrom .
MCPH1 -koder for mikrocefalin , og lidelser i dette og i ASPM er forbundet med mikrocefali. Mutasjoner i genet NBS1 som koder for nibrin kan forårsake Nijmegen bruddsyndrom , preget av mikrocefali.
Mutasjoner i EMX2 og COL4A1 er assosiert med schizencephaly , en tilstand preget av fravær av store deler av hjernehalvdelene.
Historie
I 1909 skilte Korbinian Brodmann forskjellige områder av neocortex basert på cytoarchitectural forskjell og delte hjernebarken i 52 regioner.
Rafael Lorente de Nó , student av Santiago Ramon y Cajal identifiserte mer enn 40 forskjellige typer kortikale nevroner basert på fordelingen av deres dendritter og aksoner.
Andre dyr
Hjernebarken er avledet fra pallium , en lagdelt struktur som finnes i forhjernen til alle virveldyr . Palliums grunnform er et sylindrisk lag som omslutter væskefylte ventrikler. Rundt omkretsen av sylinderen er fire soner, dorsal pallium, medial pallium, ventral pallium og lateral pallium, som antas å være homologe med henholdsvis neocortex , hippocampus , amygdala og olfactory cortex .
Inntil nylig hadde ingen motpart til hjernebarken blitt gjenkjent hos virvelløse dyr. Men en studie publisert i tidsskriftet Cell i 2010, på grunnlag av genekspresjonsprofiler, rapportert sterke affinitet mellom den cerebrale cortex og sopp legemer av ragworm Platynereis dumerilii . Sopplegemer er strukturer i hjernen til mange typer ormer og leddyr som er kjent for å spille viktige roller i læring og minne; det genetiske beviset indikerer en felles evolusjonær opprinnelse, og indikerer derfor at opprinnelsen til de tidligste forløperne i hjernebarken dateres tilbake til den tidlige prekambriske æra.
Flere bilder
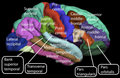
Lateral overflate av den menneskelige hjernebarken

Medial overflate av den menneskelige hjernebarken


Se også
Referanser
Eksterne linker
- hier-20 på NeuroNames
- Fargede hjerneskivebilder som inkluderer "cerebral cortex" ved BrainMaps -prosjektet
- "Den primære visuelle cortex" , Webvision: Omfattende artikkel om strukturen og funksjonen til den primære visuelle cortex.
- "Grunnleggende celletyper" , Webvision: Bilde av de grunnleggende celletypene i apen cerebral cortex.
- Cerebral Cortex - cellesentrert database
{kind=link}