
Klorofyll - Chlorophyll




Klorofyll (også klorofyl ) er et av flere beslektede grønne pigmenter som finnes i mesosomene til cyanobakterier og i kloroplastene til alger og planter . Navnet er avledet fra de greske ordene χλωρός , khloros ("lysegrønn") og φύλλον , phyllon ("blad"). Klorofyll er avgjørende for fotosyntesen , slik at planter kan absorbere energi fra lys.
Klorofyll absorberer lyset sterkest i den blå delen av det elektromagnetiske spekteret så vel som den røde delen. Motsatt er det en dårlig absorber av grønne og nesten grønne deler av spekteret. Derfor fremstår klorofyllholdige vev grønt fordi grønt lys, diffust reflektert av strukturer som cellevegger, blir mindre absorbert. To typer klorofyll finnes i fotosystemene til grønne planter: klorofyll a og b .
Historie
Klorofyll ble først isolert og navngitt av Joseph Bienaimé Caventou og Pierre Joseph Pelletier i 1817. Tilstedeværelsen av magnesium i klorofyll ble oppdaget i 1906, og var elementets første påvisning i levende vev.
Etter første arbeid utført av den tyske kjemikeren Richard Willstätter som spenner fra 1905 til 1915, ble den generelle strukturen til klorofyll a belyst av Hans Fischer i 1940. I 1960, da det meste av stereokjemien til klorofyll a var kjent, publiserte Robert Burns Woodward en total syntese av molekylet. I 1967 ble den siste gjenværende stereokjemiske belysningen fullført av Ian Fleming , og i 1990 publiserte Woodward og medforfattere en oppdatert syntese. Klorofyll f ble kunngjort for å være tilstede i cyanobakterier og andre oksygenholdige mikroorganismer som danner stromatolitter i 2010; en molekylformel på C 55 H 70 O 6 N 4 Mg og en struktur av (2- formyl ) -chlorophyll en ble utledet basert på NMR, optisk og massespektra.
Fotosyntese

Klorofyll er avgjørende for fotosyntesen , som gjør at planter kan absorbere energi fra lys .
Klorofyll molekyler er anordnet i og rundt photo som er innebygd i de thylakoid membraner i kloroplaster . I disse kompleksene tjener klorofyll tre funksjoner. Funksjonen til de aller fleste klorofyll (opptil flere hundre molekyler per fotosystem) er å absorbere lys. Etter å ha gjort det, utfører de samme sentrene sin andre funksjon: overføring av lysenergi ved hjelp av resonansoverføring til et bestemt klorofyllpar i reaksjonssenteret i fotosystemene. Dette paret påvirker den endelige funksjonen til klorofyll, ladningsseparasjon, noe som fører til biosyntese. De to godkjente fotosystemenhetene er fotosystem II og fotosystem I , som har sine egne distinkte reaksjonssentre , henholdsvis P680 og P700 . Disse sentrene er oppkalt etter bølgelengden (i nanometer ) for maksimal absorpsjon av rødtopp. Identiteten, funksjonen og spektralegenskapene til klorofylltypene i hvert fotosystem er forskjellige og bestemmes av hverandre og proteinstrukturen som omgir dem. Når de er ekstrahert fra proteinet til et løsningsmiddel (for eksempel aceton eller metanol ), kan disse klorofyllpigmentene skilles i klorofyll a og klorofyll b .
Funksjonen til reaksjonssenteret i klorofyll er å absorbere lysenergi og overføre den til andre deler av fotosystemet. Den absorberte energien til fotonet overføres til et elektron i en prosess som kalles ladningsseparasjon. Fjerning av elektronet fra klorofyllet er en oksidasjonsreaksjon. Klorofyll donerer elektronen med høy energi til en serie molekylære mellomprodukter som kalles en elektrontransportkjede . Det ladede reaksjonssenteret for klorofyll (P680 + ) reduseres deretter tilbake til grunntilstand ved å godta et elektron som er fjernet fra vann. Elektronet som reduserer P680 + kommer til slutt fra oksidasjon av vann til O 2 og H + gjennom flere mellomprodukter. Denne reaksjonen er hvordan fotosyntetiske organismer som planter produserer O 2 -gass, og er kilden til praktisk talt alt O 2 i jordens atmosfære. Fotosystem I fungerer vanligvis i serie med Fotosystem II; dermed blir P700 + av fotosystem I vanligvis redusert ettersom det aksepterer elektronet, via mange mellomprodukter i thylakoidmembranen, av elektroner som til slutt kommer fra Photosystem II. Elektronoverføringsreaksjoner i thylakoidmembranene er imidlertid komplekse, og kilden til elektroner som brukes til å redusere P700 + kan variere.
Elektronstrømmen produsert av klorofyllpigmentene i reaksjonssenteret brukes til å pumpe H + -ioner over tylakoidmembranen, og danner et kjemiosmotisk potensial som hovedsakelig brukes i produksjonen av ATP (lagret kjemisk energi) eller for å redusere NADP + til NADPH . NADPH er et universelt middel som brukes til å redusere CO 2 til sukker, så vel som andre biosyntetiske reaksjoner.
Reaksjonssenteret klorofyll -proteinkomplekser er i stand til direkte å absorbere lys og utføre ladningsseparasjonshendelser uten hjelp fra andre klorofyllpigmenter, men sannsynligheten for at det skjer under en gitt lysintensitet er liten. Således absorberer og transporterer alle de andre klorofyllene i fotosystemet og antennepigmentproteinene lysenergi til reaksjonssenteret. Foruten klorofyll a , er det andre pigmenter, kalt tilbehørspigmenter , som forekommer i disse pigment -protein antennekompleksene.
Kjemisk struktur

Klorofyller er mange i typer, men alle er definert av tilstedeværelsen av en femte ring utover de fire pyrrollignende ringene. De fleste klorofyll er klassifisert som klorofyll , som er redusert slektninger av porfyriner (finnes i hemoglobin ). De deler en felles biosyntetisk vei med porfyriner, inkludert forløperen uroporphyrinogen III . I motsetning til hemes, som har jern på midten av den porfyrin basert tetrapyrrol ring, i klorofyller sentrale magnesium- atomkoordinater med klorin , en delvis redusert porfyrin. For strukturene som er avbildet i denne artikkelen, er noen av ligandene festet til Mg 2+ sentrum utelatt for klarhet. Klorringen kan ha forskjellige sidekjeder, vanligvis inkludert en lang fytolkjede . Den mest distribuerte formen i terrestriske planter er klorofyll a .
Strukturene til klorofyller er oppsummert nedenfor:
| Klorofyll a | Klorofyll b | Klorofyll c 1 | Klorofyll c 2 | Clorophyll d | Klorofyll f | |
|---|---|---|---|---|---|---|
| Molekylær formel | C 55 H 72 O 5 N 4 mg | C 55 H 70 O 6 N 4 mg | C 35 H 30 O 5 N 4 mg | C 35 H 28 O 5 N 4 mg | C 54 H 70 O 6 N 4 mg | C 55 H 70 O 6 N 4 mg |
| C2 gruppe | -CH 3 | -CH 3 | -CH 3 | -CH 3 | -CH 3 | −CHO |
| C3 -gruppen | −CH = CH 2 | −CH = CH 2 | −CH = CH 2 | −CH = CH 2 | −CHO | −CH = CH 2 |
| C7 gruppe | -CH 3 | −CHO | -CH 3 | -CH 3 | -CH 3 | -CH 3 |
| C8 gruppe | −CH 2 CH 3 | −CH 2 CH 3 | −CH 2 CH 3 | −CH = CH 2 | −CH 2 CH 3 | −CH 2 CH 3 |
| C17 gruppe | −CH 2 CH 2 COO − Phytyl | −CH 2 CH 2 COO − Phytyl | -CH = CHCOOH | -CH = CHCOOH | −CH 2 CH 2 COO − Phytyl | −CH 2 CH 2 COO − Phytyl |
| C17 − C18 binding | Singel (klor) |
Singel (klor) |
Dobbelt (porfyrin) |
Dobbelt (porfyrin) |
Singel (klor) |
Singel (klor) |
| Hendelse | Universell | Stort sett planter | Ulike alger | Ulike alger | Cyanobakterier | Cyanobakterier |
- Strukturer av klorofyller

klorofyll a
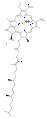
klorofyll b

klorofyll c 1

klorofyll c 2
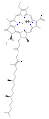
klorofyll d

klorofyll f


Måling av klorofyllinnhold

Måling av lysabsorpsjon kompliseres av løsningsmidlet som brukes til å trekke ut klorofyllet fra plantemateriale, noe som påvirker de oppnådde verdiene,
- I dietyleter har klorofyll a omtrentlig absorbansmaksimum på 430 nm og 662 nm, mens klorofyll b har omtrentlige maksima på 453 nm og 642 nm.
- Absorpsjonstoppene til klorofyll a er ved 465 nm og 665 nm. Klorofyll a fluorescerer ved 673 nm (maksimum) og 726 nm. Den maksimale molære absorpsjonskoeffisienten for klorofyll a overstiger 10 5 M −1 cm −1 , som er blant de høyeste for småmolekylære organiske forbindelser.
- I 90% aceton-vann er toppabsorberingsbølgelengdene for klorofyll a 430 nm og 664 nm; toppene for klorofyll b er 460 nm og 647 nm; toppene for klorofyll c 1 er 442 nm og 630 nm; topper for klorofyll c 2 er 444 nm og 630 nm; toppene for klorofyll d er 401 nm, 455 nm og 696 nm.
Ved å måle absorpsjonen av lys i de røde og langt røde områdene, er det mulig å estimere konsentrasjonen av klorofyll i et blad.
Forholdsfluorescensemisjon kan brukes til å måle klorofyllinnhold. Ved spennende klorofyll en fluorescens ved lavere bølgelengde, forholdet mellom klorofyllfluorescensemisjon ved705 ± 10 nm og735 ± 10 nm kan gi et lineært forhold mellom klorofyllinnhold sammenlignet med kjemisk testing. Forholdet F 735 / F 700 gitt en korrelasjonsverdi av r 2 0,96 sammenlignet med kjemiske tester i området fra 41 mg m -2 opp til 675 mg m -2 . Gitelson utviklet også en formel for direkte avlesning av klorofyllinnhold i mg m −2 . Formelen ga en pålitelig metode for å måle klorofyllinnhold fra 41 mg m −2 opp til 675 mg m −2 med en korrelasjon r 2 verdi på 0,95.
Biosyntese
I noen planter stammer klorofyll fra glutamat og syntetiseres langs en forgrenet biosyntetisk vei som deles med hem og siroheme . Klorofyllsyntase er enzymet som fullfører biosyntesen av klorofyll a ved å katalysere reaksjonen EC 2.5.1.62
- klorofyllid a + fytyldifosfat klorofyll a + difosfat

Dette danner en ester av karboksylsyre-gruppen i chlorophyllide en med den 20-karbon- diterpen alkohol fytol . Klorofyll b er laget av det samme enzymet som virker på klorofyllid b .
I Angiosperm- planter er de senere trinnene i den biosyntetiske banen lysavhengige, og slike planter er bleke ( etiolert ) hvis de vokser i mørke. Ikke-vaskulære planter og grønne alger har et ekstra lysuavhengig enzym og vokser grønt selv i mørket.
Klorofyll i seg selv er bundet til proteiner og kan overføre den absorberte energien i ønsket retning. Protochlorophyllide , et av de biosyntetiske mellomproduktene, forekommer hovedsakelig i fri form og fungerer under lysforhold som en lysfølsomhet og danner svært giftige frie radikaler . Derfor trenger planter en effektiv mekanisme for å regulere mengden av denne klorofyllforløperen. I angiospermer gjøres dette på trinnet med aminolevulinsyre (ALA), en av mellomforbindelsene i biosyntesebanen. Planter som mates av ALA akkumulerer høye og giftige nivåer av protochlorophyllide; det samme gjør mutantene med et skadet reguleringssystem.
Aldring og klorofyllsyklusen
Prosessen med plantens aldring involverer nedbrytning av klorofyll: for eksempel enzymet klorofyllase ( EC 3.1.1.14 ) hydrolyserer fytylsidekjeden for å reversere reaksjonen der klorofyll blir biosyntetisert fra klorofyllid a eller b . Siden klorofyllid a kan omdannes til klorofyllid b og sistnevnte kan omestres til klorofyll b , tillater disse prosessene å sykle mellom klorofyll a og b . Videre kan klorofyll b reduseres direkte (via 7 1 -hydroksyklorofyll a ) tilbake til klorofyll a og fullføre syklusen. I senere stadier av aldring blir klorofyllider omdannet til en gruppe fargeløse tetrapyrroler kjent som ikke -fluorescerende klorofyllkatabolitter (NCC) med den generelle strukturen:

Disse forbindelsene har også blitt identifisert i modning av frukt, og de gir karakteristiske høstfarger til løvfellende planter.
Defekte miljøer kan forårsake klorose
Klorose er en tilstand der blader produserer utilstrekkelig klorofyll og gjør dem gule. Klorose kan skyldes næringsstoffmangel på jern - kalt jernklorose - eller mangel på magnesium eller nitrogen . Jordens pH spiller noen ganger en rolle i næringsårsaket klorose; mange planter er tilpasset til å vokse i jord med spesifikke pH -nivåer, og deres evne til å ta opp næringsstoffer fra jorda kan være avhengig av dette. Klorose kan også skyldes patogener, inkludert virus, bakterier og soppinfeksjoner eller saftsugende insekter.
Komplementær lysabsorbering av antocyaniner
.PNG)
Anthocyaniner er andre plantepigmenter . Absorbansmønsteret som er ansvarlig for den røde fargen på antocyaniner kan være komplementær med grønt klorofyll i fotosyntetisk aktive vev som unge Quercus coccifera -blader . Det kan beskytte bladene mot angrep fra planteeter som kan tiltrekkes av grønn farge.
Fordeling
Klorofyllkartene viser milligram klorofyll per kubikkmeter sjøvann hver måned. Steder der klorofyllmengdene var svært lave, noe som indikerer svært lave fytoplankton , er blå. Steder hvor klorofyllkonsentrasjonene var høye, noe som betyr at mange planteplankton vokste, er gule. Observasjonene kommer fra Moderate Resolution Imaging Spectroradiometer (MODIS) på NASAs Aqua -satellitt. Land er mørkegrått, og steder der MODIS ikke kunne samle inn data på grunn av havis, polarmørke eller skyer er lysegrå. De høyeste klorofyllkonsentrasjonene, hvor bittesmå overflateboende havplanter trives , er i kalde polare farvann eller på steder der havstrømmer bringer kaldt vann til overflaten, for eksempel rundt ekvator og langs kysten av kontinenter. Det er ikke det kalde vannet i seg selv som stimulerer planteplanktonet. I stedet er de kule temperaturene ofte et tegn på at vannet har hevet seg opp til overflaten fra dypere i havet og bærer næringsstoffer som har bygget seg opp over tid. I polvann akkumuleres næringsstoffer i overflatevannet i de mørke vintermånedene når planter ikke kan vokse. Når sollyset kommer tilbake om våren og sommeren, blomstrer plantene i høye konsentrasjoner.
Kulinarisk bruk
Syntetisk klorofyll er registrert som et tilsetningsstoff i tilsetningsstoff, og E -nummeret er E140 . Kokker bruker klorofyll for å farge en rekke matvarer og drikkevarer grønne, for eksempel pasta og brennevin. Absint får sin grønne farge naturlig fra klorofyllet som ble introdusert gjennom det store utvalget av urter som ble brukt i produksjonen. Klorofyll er ikke løselig i vann, og det blandes først med en liten mengde vegetabilsk olje for å oppnå ønsket løsning .
Biologisk bruk
En studie fra 2002 fant at "blader utsatt for sterkt lys inneholdt nedbrutte store antenneproteiner , i motsetning til de som er holdt i mørket, noe som er i samsvar med studier om belysning av isolerte proteiner ". Dette fremsto for forfatterne som støtte for hypotesen om at " aktive oksygenarter spiller en rolle in vivo " i planters kortsiktige oppførsel.
Se også
- Bakterioklorofyll , beslektede forbindelser i fototrofe bakterier
- Klorofyllin , et halvsyntetisk derivat av klorofyll
- Maksimal dyp klorofyll
- Grow light , en lampe som fremmer fotosyntesen
- Klorofyllfluorescens , for å måle plantestress