
Energistrøm (økologi) - Energy flow (ecology)
Energistrøm er energistrømmen gjennom levende ting i et økosystem. Alle levende organismer kan organiseres i produsenter og forbrukere, og disse produsentene og forbrukerne kan videre organiseres i en næringskjede . Hvert av nivåene i næringskjeden er et trofisk nivå. For mer effektivt å vise mengden organismer på hvert trofisk nivå, blir disse næringskjedene deretter organisert i trofiske pyramider. Pilene i næringskjeden viser at energistrømmen er enveis, pilens hode viser retning energien beveger seg inn, og at energien går tapt som varme i hvert trinn underveis.
Den ensrettede energistrømmen og det påfølgende energitapet når det beveger seg opp i matvevet, er mønstre i energistrømmen som styres av termodynamikk , som er begrepet energibytte mellom systemer. Trofisk dynamikk forholder seg til termodynamikk fordi den omhandler overføring og transformasjon av energi (som kommer eksternt fra solen via solstråling) til og blant organismer.

Energi og karbonsyklus

100 × 10 15 gram karbon/år fikset av fotosyntetiske organismer, noe som tilsvarer 4 × 10 18 kJ/år = 4 × 10 21 J/år fri energi.Celleaspirasjon er omvendt reaksjon, hvor energi fra planter og oksygen tas opp og frigjøres når karbondioksid og vann avgis. Koldioksid og vann som produseres kan resirkuleres tilbake til planter.
Det første trinnet i energien er fotosyntese , der vann og karbondioksid fra luften tas opp med energi fra solen og omdannes til oksygen og glukose. Cellepusting er omvendt reaksjon, der oksygen og sukker tas opp og frigjør energi når de omdannes til karbondioksid og vann. Kullsyre og vann produsert ved respirasjon kan resirkuleres tilbake til planter.
Energitap kan måles enten ved effektivitet (hvor mye energi går til neste nivå), eller ved biomasse (hvor mye levende materiale som finnes på disse nivåene på et tidspunkt, målt ved stående avling). Av all netto primær produktivitet på produsentens trofiske nivå går generelt bare ti prosent til neste nivå, primærforbrukerne, deretter går bare ti prosent av de ti prosentene videre til neste trofiske nivå, og så videre opp i matpyramiden . Økologisk effektivitet kan være alt fra 5% til 20% avhengig av hvor effektivt eller ineffektivt det økosystemet er. Denne reduksjonen i effektivitet skjer fordi organismer må utføre cellulær respirasjon for å overleve, og energi går tapt som varme når mobil respirasjon utføres. Det er også derfor det er færre tertiære forbrukere enn det er produsenter.
Primærproduksjon
En produsent er enhver organisme som utfører fotosyntese. Produsenter er viktige fordi de omdanner energi fra solen til en lagringsbar og brukbar kjemisk form for energi, glukose, så vel som oksygen. Produsentene kan bruke energien lagret i glukose og oksygen til å utføre cellulær respirasjon. Eller, hvis produsenten blir konsumert av planteetere i det neste trofiske nivået, blir noe av energien ført videre til pyramiden. Glukosen som er lagret hos produsentene, fungerer som mat for forbrukere, og det er bare gjennom produsenter at forbrukerne får tilgang til solens energi. Noen eksempler på primærprodusenter er alger, moser og andre planter som gress, trær og busker.
Kjemosyntetiske bakterier utfører en prosess som ligner fotosyntese, men i stedet for energi fra solen bruker de energi lagret i kjemikalier som hydrogensulfid og oksygen. Denne prosessen, referert til som kjemosyntese , skjer vanligvis dypt i havet ved hydrotermiske ventiler som produserer varme og kjemikalier som hydrogen, hydrogensulfid og metan. Kjemosyntetiske bakterier kan bruke energien i bindingene til hydrogensulfid og oksygen for å omdanne karbondioksid til glukose, og frigjøre vann og svovel i prosessen. Organismer som forbruker de kjemosyntetiske bakteriene kan ta opp glukosen og bruke oksygen til å utføre cellulær respirasjon, på samme måte som planteetere som bruker produsenter.
En av faktorene som styrer primærproduksjonen er mengden energi som kommer inn i produsentene, som kan måles ved hjelp av produktivitet. Bare en prosent av solenergien kommer inn i produsenten, resten spretter av eller beveger seg gjennom. Brutto primærproduktivitet er mengden energi produsenten faktisk får. Vanligvis går 60 % av energien som kommer inn i produsenten til produsentens eget åndedrett. Netto primær produktivitet er mengden planten får etter at mengden den brukte til mobil respirasjon er trukket fra. En annen faktor som styrer primærproduksjonen er organiske/uorganiske næringsnivåer i vannet eller jorda som produsenten lever i.
Sekundær produksjon
Sekundær produksjon er bruk av energi lagret i anlegg som omdannes av forbrukere til deres egen biomasse. Ulike økosystemer har forskjellige nivåer av forbrukere, alle ender med en toppforbruker. Mest energi lagres i planter og oksygen, og når forbrukerne spiser disse plantene, tar de opp en liten mengde energi. Denne energien i planteetere og altetende blir deretter konsumert av kjøttetere. Det er også en stor mengde energi som er i primærproduksjon og ender med å bli avfall eller søppel, referert til som detritus. Den skadelige næringskjeden inneholder en stor mengde mikrober, makroinvertebrater, meiofauna, sopp og bakterier. Disse organismene blir konsumert av altetende og rovdyr og står for en stor mengde sekundærproduksjon. Sekundære forbrukere kan variere mye i hvor effektive de er i forbruket. Effektiviteten av energi som overføres til forbrukere anslås å være rundt 10%. Energistrømmen gjennom forbrukere er forskjellig i vann- og terrestriske miljøer.
Sekundær produksjon i vannmiljøer
Heterotrofer bidrar til sekundærproduksjon, og det er avhengig av primærproduktivitet og netto primære produkter. Sekundær produksjon er energien som planteetere og nedbrytere bruker, og dermed er sekundær produktivitet avhengig av primær produktivitet. Hovedsakere og nedbrytere bruker først og fremst alt karbon fra to hovedkilder i akvatiske økosystemer. De to typene viktige karbon fra organiske kilder er autokton og allokton. Autoktonisk karbon kommer fra økosystemet og inkluderer vannplanter, alger og planteplankton. Allochthonous carbon utenfra økosystemet er stort sett dødt organisk materiale fra det terrestriske økosystemet som kommer inn i vannet. I bekkøkosystemer kan omtrent 66% av den årlige energitilførselen vaskes nedstrøms. Den gjenværende mengden forbrukes og går tapt som varme.
Sekundær produksjon i terrestriske miljøer
Sekundær produksjon er ofte beskrevet i form av trofiske nivåer , og selv om dette kan være nyttig for å forklare relasjoner, legger det for mye vekt på de sjeldnere interaksjonene. Forbrukere spiser ofte på flere trofiske nivåer. Energi overført over det tredje trofiske nivået er relativt uviktig. Assimileringseffektiviteten kan uttrykkes ved mengden mat forbrukeren har spist, hvor mye forbrukeren assimilerer og hva som blir utvist som avføring eller urin. Mens en del av energien brukes til respirasjon, går en annen del av energien til biomasse hos forbrukeren. Det er to store næringskjeder: Den primære næringskjeden er energien som kommer fra autotrofer og sendes videre til forbrukerne; og den andre store næringskjeden er når rovdyr spiser planteetere eller nedbrytere som bruker den autotrofiske energien. Forbrukerne er delt inn i hovedforbrukere, sekundære forbrukere og tertiære forbrukere. Rovdyr har en mye høyere assimilering av energi, omtrent 80% og planteetere har en mye lavere effektivitet på omtrent 20 til 50%. Energi i et system kan påvirkes av dyreutvandring/innvandring. Organismenes bevegelser er signifikante i terrestriske økosystemer. Energisk forbruk av planteetere i terrestriske økosystemer har et lavt område på ~ 3-7%. Energistrømmen er lik i mange terrestriske miljøer. Svingningen i mengden netto primært produkt som forbrukes av planteetere er generelt lav. Dette står i stor kontrast til vannmiljøer i innsjøer og dammer der beiter har et mye høyere forbruk på rundt ~ 33%. Ectotherms og endotherms har svært forskjellige assimileringseffektiviteter.
Detritivores
Detritivores bruker organisk materiale som brytes ned og blir igjen konsumert av kjøttetere. Rovdyrproduktivitet er korrelert med byttedyrproduktivitet. Dette bekrefter at den primære produktiviteten i økosystemer påvirker all produktivitet som følger.
Detritus er en stor del av organisk materiale i økosystemer. Organisk materiale i tempererte skoger består for det meste av døde planter, omtrent 62%.
I et akvatisk økosystem blir bladmateriale som faller i bekker vått og begynner å igle organisk materiale, det skjer ganske raskt og vil tiltrekke seg mikrober og virvelløse dyr. Bladene kan brytes ned i store biter som kalles naturlig partikkelformig organisk materiale (CPOM). CPOM koloniseres raskt av mikrober. Meiofauna er ekstremt viktig for sekundærproduksjon i bekkøkosystemer . Mikrober som bryter ned og koloniserer på dette bladstoffet er svært viktige for detritovorene. Detritovorene gjør bladmaterialet mer spiselig ved å frigjøre forbindelser fra vevet; det hjelper til slutt med å myke dem. Ettersom bladene henfaller, vil nitrogen avta siden cellulose og lignin i bladene er vanskelig å bryte ned. Dermed bringer de koloniserende mikrober inn nitrogen for å hjelpe til med nedbrytningen. Nedbrytning av blad kan avhenge av innledende nitrogeninnhold, sesong og treslag. Treslagene kan ha variasjon når bladene faller. Dermed skjer nedbrytning av blader på forskjellige tidspunkter, som kalles en mosaikk av mikrobielle populasjoner.
Arteffekt og mangfold i et økosystem kan analyseres gjennom ytelse og effektivitet. I tillegg kan sekundærproduksjon i bekker bli sterkt påvirket av detritus som faller ned i bekkene; produksjonen av bunndyrbiomasse og overflod reduserte ytterligere 47–50% under en studie av fjerning og utestenging av søppel.
Energistrøm på tvers av økosystemer
Forskning har vist at primærprodusenter fikser karbon med lignende hastigheter på tvers av økosystemer. Når karbon er blitt introdusert i et system som en levedyktig energikilde, varierer mekanismene som styrer strømmen av energi til høyere trofiske nivåer på tvers av økosystemer. Blant akvatiske og terrestriske økosystemer er det identifisert mønstre som kan forklare denne variasjonen og har blitt delt inn i to hovedveier for kontroll: ovenfra og ned og ned-opp. De virkende mekanismene i hver vei regulerer til slutt strukturen i fellesskap og trofisk nivå i et økosystem i ulik grad. Bottom-up-kontroller involverer mekanismer som er basert på ressurskvalitet og tilgjengelighet, som styrer primærproduktivitet og den påfølgende strømmen av energi og biomasse til høyere trofiske nivåer. Top-down kontroller involverer mekanismer som er basert på forbruk av forbrukere. Disse mekanismene styrer hastigheten på energioverføring fra et trofisk nivå til et annet ettersom planteetere eller rovdyr lever av lavere trofiske nivåer.
Akvatiske vs terrestriske økosystemer
Mye variasjon i energistrømmen finnes innenfor hver type økosystem, noe som skaper en utfordring i å identifisere variasjon mellom økosystemtyper. I generell forstand er energistrømmen en funksjon av primær produktivitet med temperatur, vanntilgjengelighet og lystilgjengelighet. For eksempel, blant akvatiske økosystemer, finnes vanligvis høyere produksjonshastigheter i store elver og grunne innsjøer enn i dype innsjøer og klare vann. Blant terrestriske økosystemer har myrer, sump og tropiske regnskoger de høyeste primære produksjonshastighetene, mens tundra og alpine økosystemer har de laveste primære produksjonshastighetene. Forholdet mellom primærproduksjon og miljøforhold har bidratt til å ta hensyn til variasjon innenfor økosystemtyper, slik at økologer kan demonstrere at energi flyter mer effektivt gjennom akvatiske økosystemer enn terrestriske økosystemer på grunn av de forskjellige bunn-opp og topp-ned-kontrollene i spillet.
Opp ned
Styrken til bunn-opp-kontroller på energistrømmen bestemmes av ernæringskvaliteten, størrelsen og vekstraten til primærprodusenter i et økosystem. Fotosyntetisk materiale er vanligvis rikt på nitrogen (N) og fosfor (P) og supplerer den høye etterspørselen etter planteetere etter N og P i alle økosystemer. Akvatisk primærproduksjon domineres av små, encellede fytoplankton som hovedsakelig består av fotosyntetisk materiale, som gir en effektiv kilde til disse næringsstoffene for planteetere. I kontrast inneholder flercellede terrestriske planter mange store støttende cellulosestrukturer med høyt karbon, men lav næringsverdi. På grunn av denne strukturelle forskjellen har akvatiske primærprodusenter mindre biomasse per fotosyntetisk vev lagret i det akvatiske økosystemet enn i skog og gressletter i terrestriske økosystemer. Denne lave biomassen i forhold til fotosyntetisk materiale i akvatiske økosystemer gir mer effektiv omsetningshastighet sammenlignet med terrestriske økosystemer. Ettersom planteplankton forbrukes av planteetere, erstatter deres forbedrede vekst og reproduksjonshastighet tilstrekkelig tapt biomasse og støtter i tillegg til næringstett kvalitet større sekundærproduksjon.
Ytterligere faktorer som påvirker primærproduksjonen inkluderer tilførsler av N og P, som forekommer i større omfang i akvatiske økosystemer. Disse næringsstoffene er viktige for å stimulere plantevekst, og når de overføres til høyere trofiske nivåer, stimulerer de forbrukerens biomasse og veksthastighet. Hvis noen av disse næringsstoffene mangler, kan de begrense den totale primærproduksjonen. Innen innsjøer pleier P å være det største begrensende næringsstoffet, mens både N og P begrenser primærproduksjonen i elver. På grunn av disse begrensende effektene kan tilførsel av næringsstoffer potensielt lindre begrensningene på netto primærproduksjon av et akvatisk økosystem. Allochthonous materiale vasket inn i et akvatisk økosystem introduserer N og P samt energi i form av karbonmolekyler som lett tas opp av primærprodusenter. Større tilførsler og økte næringsstoffkonsentrasjoner støtter større netto primærproduksjonshastigheter, noe som igjen støtter større sekundærproduksjon.
Ovenfra og ned
Øverst ned-ned-mekanismer utøver større kontroll på primære produsenter i vann på grunn av forbrukerrullen i et vannmatnett. Blant forbrukerne kan planteetere formidle virkningen av trofiske kaskader ved å bygge bro mellom energistrømmen fra primærprodusenter til rovdyr på høyere trofiske nivåer. På tvers av økosystemer er det en konsekvent sammenheng mellom plantevekstvekst og produsentens ernæringsmessige kvalitet. I akvatiske økosystemer blir primærprodusentene imidlertid konsumert av planteetere i en hastighet som er fire ganger større enn i terrestriske økosystemer. Selv om dette emnet er sterkt omdiskutert, har forskere tilskrevet skillet i planteetende bekjempelse til flere teorier, inkludert størrelsesforhold mellom produsent og forbruker og planteetende selektivitet.
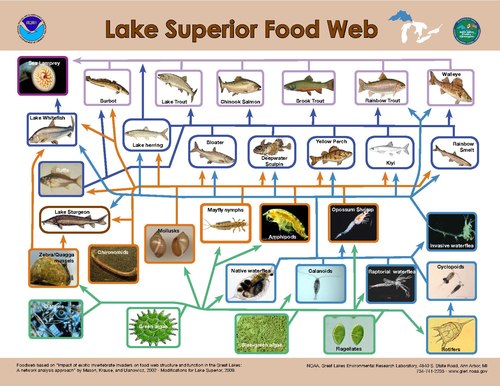
Modellering av topp-ned-kontroller på primærprodusenter antyder at den største kontrollen med energistrømmen skjer når størrelsesforholdet mellom forbruker og primærprodusent er det høyeste. Størrelsesfordelingen av organismer som finnes innenfor et enkelt trofisk nivå i akvatiske systemer er mye smalere enn for terrestriske systemer. På land varierer forbrukerstørrelsen fra mindre enn planten den forbruker, for eksempel et insekt, til betydelig større, for eksempel en hovdyr , mens i akvatiske systemer varierer forbrukerkroppens størrelse innenfor et trofisk nivå mye mindre og er sterkt korrelert med trofisk posisjon. Som et resultat er størrelsesforskjellen mellom produsenter og forbrukere konsekvent større i vannmiljøer enn på land, noe som resulterer i sterkere planteetende kontroll over akvatiske primærprodusenter.
Gressetere kan potensielt kontrollere skjebnen til organisk materiale når det sykles gjennom matvevet. Gressetere pleier å velge næringsrike planter mens de unngår planter med strukturelle forsvarsmekanismer . I likhet med støttestrukturer består forsvarsstrukturer av næringsfattig, høy karbon cellulose. Tilgang til næringsrike matkilder øker metabolismen av planteetere og energibehov, noe som fører til større fjerning av primærprodusenter. I akvatiske økosystemer er planteplankton svært næringsrikt og mangler generelt forsvarsmekanismer. Dette resulterer i større topp-ned-kontroll fordi forbrukte plantestoffer raskt slippes tilbake i systemet som labilt organisk avfall . I terrestriske økosystemer er primærprodusenter mindre ernæringsmessig tette og er mer sannsynlig å inneholde forsvarsstrukturer. Fordi planteetere foretrekker ernæringsmessig tette planter og unngår planter eller plantedeler med forsvarsstrukturer, blir en større mengde plantemateriale igjen uforbruket i økosystemet. Unngåelse av planteetere av lavkvalitets plantemateriale kan være grunnen til at terrestriske systemer viser svakere topp-ned-kontroll på energistrømmen.
Se også
Referanser
Videre lesning
- Podolinsky S (2004). "Sosialisme og enhet i fysiske krefter". Organisasjon og miljø . 17 (1): 61–75. doi : 10.1177/1086026603262092 . S2CID 144332347 .
- Weiner DR (2000). Modeller av natur: Økologi, bevaring og kulturrevolusjon i Sovjet -Russland . USA: University of Pittsburgh Press .