
Epitelioidcelle - Epithelioid cell

Ifølge et vanlig synspunkt er epitelioider (også kalt epitelioid histiocytter ) derivater av aktiverte makrofager som ligner epitelceller .
Struktur og funksjon

Strukturelt er epitelioidceller (når de undersøkes ved lysmikroskopi etter farget med hematoksylin og eosin), forlenget med fint granulært, blekt eosinofilt (rosa) cytoplasma og sentrale, eggformede kjerner (ovale eller langstrakte), som er mindre tette enn for en lymfocytt . De har utydelig form og ser ut til å smelte sammen til hverandre og danne aggregater kjent som gigantiske celler. Når de undersøkes ved overføringselektronmikroskopi i epitelceller i feltet av Golgi -lamellkomplekset, teipes ikke bare sonerte, men også slanke vesikler med tett senter, og også mange (mer enn 100) store granuler med diametere opp til 340 nm og med granulat matrise mer lys enn i makrofaggranulater, noen ganger med perigranulær glorie. “Det mest fremtredende trekket ved disse cellene er det enorme Golgi -området; opptil 6 individuelle stabler med Golgi cisternae kan være tilstede, så vel som noen få børstehårslakkede og mange glatte vesikler ”. Epitelioidceller har tett interdigiterte cellemembraner i glidelåslignende matriser som forbinder tilstøtende celler. Disse cellene er sentrale i dannelsen av granulomer, som er forbundet med mange alvorlige sykdommer. I granulomer utfører epitelioidceller avgrensningsfunksjonene.
Særegenheter ved cytoskjelettet

Det er vist at cytoskjelettet av epitelceller dannet av filamenter skiller seg vesentlig fra makrofagcytoskjelettet. En stor økning i antall filamenter skjer i disse cellene, hvor filamenter (90 til 100 A) omgir cytocentrum som en særegen ringformet bunt som ofte forgrener seg til cytoplasma. På grunn av slike cytoskjeletter viser sammenhengende epitelceller utførlige cytoplasmatiske interdigitasjoner. Ved å bruke metodene for hurtig = fryse og fryse-substitusjon (hurtig frysing, penetrerende etsing og frysesubstitusjon) har det blitt vist at organisasjonene tredimensjonale metastrukturkitoskelett av epitelioidcellene, dannet i fokus av granulomatøs betennelse , mer kompatibel med cytoskjelett karakteristisk for typisk epitelcelle enn for cytoskjelett av aktive og bevegelige makrofager. Det er vist at de tette banene av mellomliggende filamenter, bundet med kjerner, mitokondrioner og andre organeller, overvåkes overalt i cytoplasma av epitelioidceller. Noen fascicles av aktiniske filamenter ble plassert i filopodiums nedenfor enn membraner i cellene. Nøyaktig interdigital tredobling av cellemembraner mellom grensesnitt -epitelioidceller ble tydelig demonstrert. Interdigitale filopodier ble identifisert. Den karakteristiske indikasjonen på epitelioidceller er deres aggregering med formasjonstette interdigitale triplinger som en "lyn" som fester, som tilsynelatende kan ha det viktige for dannelsen av en tett sone for avgrensning av kroppen fra patogenet under dannelsen av epitelioidcelle granulomer.
Immunologisk fenotype og immunologiske markører
Ved bruk av antistoffer mot RFD9-, RFD7- og HLA-DR-antigenene ble det funnet at alle epitelioidceller har en immunologisk fenotype RFD9+/RFD7-/HLA-DR+. En serie monoklonale antistoffer IHY-1, IHY-2, IHY-3 ble oppnådd, som kan brukes til å nøyaktig identifisere epitelceller dannet i etiologisk forskjellige former for granulomatøs betennelse. IHY-1-antistoff reagerer med epitelceller i sarkoidgranulomer så vel som med epitelceller av forskjellige granulomatøse sykdommer, inkludert tuberkulose. IHY-2 og IHY-3 l antistoff, monoklonalt antistoff, reagerer med epitelioidceller ved sarkoidose, men ikke i tuberkulose .
Klinisk signifikans
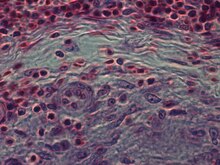
Epitelioidceller er en vesentlig egenskap ved epitelioidcellegranulomer . Epitelioidcellegranulom kan defineres som spesifikk og strukturelt organisert samling av epitelioidceller, makrofager, lymfocytter og dendritiske celler. Fremmedlegeme-granulomer kan betraktes som en organisert samling av makrofager, inkludert bare samlinger av gigantiske celler som omgir inerte stoffer som suturmateriale-de såkalte "ikke-immune granulomer." Granulomdannelse er assosiert med patogener som har lært å unnslippe vertens immunsystem på forskjellige måter som å motstå fagocytose og drap i makrofager. Ufordøyelighet av materie ved makrofager er et vanlig trekk ved granulomatøs betennelse. Granulomer prøver å fjerne disse organismer og forhindre videre vekst og spredning. Historisk utbredte og ødeleggende sykdommer som tuberkulose, spedalskhet og syfilis er granulomatiske tilstander. Granulomdannelse er også funksjonen til mange flere samtidige forhold, som soppinfeksjoner, sarkoidose og Crohns sykdom .
Vitenskapelig forskningshistorie
Den første omtale av epitelioider som en spesifikk celleform skjedde på 1800-tallet i verk av Koch R og Cornil J, som trodde at leukocyttene var opphavsmennene til epitelcellene til tuberkulosen. I forsøk på kaniner viste Yersin A (1888) og Borrel A (1893) at epitelioidceller dannes fra mononukleære blodceller i blod. Hovedmønstrene for dannelse av epitelioider ble først beskrevet i første halvdel av 1900 -tallet av Lewis M (1925). Denne forskeren viste at blodmonocytter i cellekulturer av leukocytter av blandede blod av aviær (hentet fra voksne fugler så vel som fra embryoer i forskjellige aldre), mus og mennesker, når de dyrkes in vitro , transformeres til typiske makrofager og epitelioidceller, fulgte ved dannelse av gigantiske flerkjernede celler. Dannelsen av celler av epitelioidtype ble notert av Lewis M på 2.-3. Dag av dyrking av leukocytter. Senere i en studie av en lignende plan, viste Jerry S og Weiss L (1966), når de brukte kulturer av blandede blodleukocytter av kylling (atskilt fra hjerteblod fra Rhode Island Red) og elektronmikroskopi, at transformasjonen av kyllingmonocytter i epitelioidceller begynner i kultur på 3-4 dager og slutter på 5-6 dager. Siden alle tidligere forskere har indikert at epitelioidceller dannes fra monocytter, og monocytter og makrofager ble kombinert til et enkelt mononukleært fagocyttsystem , Van Furth et al. (1972), med henvisning til arbeidet til Sutton J og Weiss L (1966), tilskrev formelt epitelioidceller det mononukleære fagocyttsystemet. Imidlertid spesifiserte de ikke nøyaktig fra hvilke celler i det mononukleære fagocyttsystemet epitelioidceller stammer fra. Samtidig kom de med en veldig forsiktig konklusjon om at «Epitelioidcellene som forekommer i disse lesjonene også stammer fra monocytter eller makrofager». Adams D (1976), og trodde at epitelioidceller er det siste stadiet av celledifferensiering av cellene i det mononukleære fagocyttsystemet, formulerte begrepet cytomorphogeesis av epitelioide celler ifølge hvilke epitelioidceller anses som et derivat av en aktivert makrofag (som er fortsatt i besittelse av de fleste forskere for tiden). Den er basert på antagelsen om at "stimulering av makrofager modnes videre til umodne epitelceller og ultimatelv hos modne". Rhee et al. (1979) i eksperimenter på rotter, ved bruk av metoden for elektronmikroskopi, viste at en av de viktigste cytomorfologiske egenskapene til epitelioide celler som skiller disse cellene fra makrofager, er tilstedeværelsen av karakteristiske spesifikke granulater i dem, som de kalte epitelioide cellekorn. Basert på egne data, støttet de konseptet med epitelioide cellers cytomorfogenese der epitelioidceller blir sett på som et derivat av en aktivert makrofag. Senere foreslo Turk JL og Narayanan RB (1982) to typer epitelioidceller i studien : "vesikulære" og "sekretoriske" epitelceller. Det antydes at "vesikulære epitelioidceller kan utvikle seg fra" sekretoriske "epitelceller ved en degenerasjonsprosess. På jakt etter immunologiske mekanismer som påvirker dannelsen av epitelceller, fikk Cipriano et al. (2003) data som indikerer mulig påvirkning av IL- 4 om dannelsen av en fenotype i makrofager som ligner fenotypen til epitelioidceller. Imidlertid passer ikke alle resultatene av undersøkelser om studiet av lovene og mekanismene for cytomorfogenese av epitelioidceller i begrepet opprinnelse til epitelioide celler fra makrofager. Deimann J og Fahimi H (1980) viste at epitelioidceller i granulomer, indusert i rottelever ved injeksjon av glukan, beta-1,30-polyglukose, dannes ikke fra Kupffer-celler -modne differensierte makrofager, men fra blod monocytter. De Vos et al. (1990) innhentet dataene som tillot dem å antyde at i granulomatøse betennelsesfoci, i granulomatøs lymfadenitt , epitelioide celler dannes ikke fra differensierte makrofager, men fra såkalte plasmacytoidmonocytter (har likheter med plasmacytter). Dette understøttes ytterligere av de ultrastrukturelle likhetene mellom plasmacytoidmonocytter og epitelioidceller. Den nåværende ultrakonstruksjonelle og immunoelektronmikroskopiske studien av epitelioidcellegranulomer ga ytterligere argumenter til fordel for denne hypotesen. Arkhipov S (1997, 2012) ved bruk av kulturer av peritoneale celler, blodleukocytter og benmargsceller hos mus, viste at makrofager og epitelioidceller dannes fra forskjellige typer monocytter. Det har blitt vist at epitelioider bare dannes av monocytter av plasmocytoid type, har navngitt pre-epitelioide celler, og omgått differensieringstrinnet til makrofager. Det har blitt vist at ved kronisk betennelse øker antallet pre-epitelioide monocytiske celler, begått i epitelioid celledifferensiering, i fokus for betennelse, i blod og benmarg. ved bruk av mus Innavlet linjer , i motsetning til mottakelighet for Mycobacterium tuberculosis , ble det vist at antallet pre-epitelioide monocytiske celler, dannet ved kronisk betennelse, er genetisk bestemt. De oppnådde resultatene viste at morfogenesen av epitelioide cellegranulomer kan bestemmes av det forskjellige utgangsgenetisk bestemte nivået i et basseng av pre-epitelioide celler av monocytoid type, deres flyt i midten av en granulomatøs betennelse, intensivere dem differensiering til epitelioidceller, omgå fase av differensiering til makrofager, og deres endomitotiske aktivitet.
Se også
Referanser
Eksterne linker
- Epitelioid+celler ved US National Library of Medicine Medical Subject Headings (MeSH)
Denne cellebiologiartikkelen er en stubbe . Du kan hjelpe Wikipedia ved å utvide den . |