Heterokont - Heterokont
| Heterokont | |
|---|---|

|
|
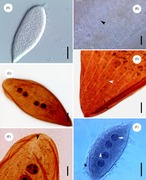
| Ochromonas sp. ( Chrysophyceae ), med to ulik (heterokont) flagella. Mastigonemer ikke representert. | |
|
Vitenskapelig klassifisering |
|
| Clade : | SAR |
| Krenkelse: |
Heterokonta Cavalier-Smith , 1986 |
| Typiske klasser | |
|
Fargede grupper ( algelignende )
Fargeløse grupper |
|
| Synonymer | |
|
|
Heterokonts er en gruppe protister (formelt referert til som Heterokonta , Heterokontae eller Heterokontophyta ). Gruppen er en hovedlinje av eukaryoter . De fleste er alger , alt fra den gigantiske flercellede taren til de encellede kiselalger , som er en hovedkomponent i plankton . Andre bemerkelsesverdige medlemmer av Stramenopiles inkluderer (generelt) parasittiske oomycetes , inkludert Phytophthora , som forårsaket Irlands store hungersnød , og Pythium , som forårsaker frøråtne og demping.
Navnet "heterokont" refererer til typen motilt livssyklusstadium, der de flagellerte cellene har to ulikt arrangerte flageller (se zoospore ).
Historie
I 1899 opprettet Alexander Luther "Heterokontae" for noen alger med ulik flagella, i dag kalt Xanthophyceae . Senere ville noen forfattere (f.eks. Copeland, 1956) inkludere andre grupper i Heterokonta og utvide sansen. Begrepet fortsetter å bli brukt på forskjellige måter, noe som fører til at Heterokontophyta også brukes på phylum Ochrophyta . Begrepet 'Stramenopile' ble introdusert i 1989 av Patterson for å overvinne uklarheter som hadde (og fortsetter å bli) utviklet ved bruk av begrepet 'heterokont'. Følgelig kan heterokoner refereres til som stramenopiler.
Begrepet 'heterokont' dukket først opp i sammenheng med fykologi fra 1800 -tallet. Over tid har anvendelsesområdet endret seg; spesielt når det på 1970 -tallet som ultrastrukturerte studier avslørte større mangfold blant alger med kromoplaster (= klorofyll a og c) enn det som tidligere var blitt anerkjent. Samtidig erstattet et protistologisk perspektiv det 19. århundre basert på inndelingen av encellede eukaryoter langs upassende botaniske/zoologiske linjer. En konsekvens var at en rekke heterotrofiske organismer, hvorav mange ikke tidligere hadde blitt betraktet som 'heterokont', ble sett på som relatert til 'kjerneheterokontene' (dvs. de som har fremre flagell med stive hår). Ny anerkjente slektninger inkluderte parasittiske opaliner, proteromonader og actinophryid heliozoa. De sluttet seg til andre heterotrofiske protister, for eksempel bicosoecider, labyrinthulids og oomycete sopp, som ble inkludert av noen som heterokonter og ekskludert av andre. I stedet for å fortsette å bruke et navn hvis betydning hadde endret seg over tid og derfor var tvetydig, ble navnet 'stramenopile' introdusert for å referere til protestenes klader som hadde trepartige stive (vanligvis flagellære) hår og alle deres etterkommere. Molekylære studier bekrefter at genene som koder for proteinene i disse hårene er eksklusive for stramenopiler. Ettersom begrepet 'Stramenopile' er basert på en antatt apomorfi, er det stabilt og robust selv når sammensetningen endres. Det er en utbredt antagelse, som her, at begrepene 'stramenopil' og 'heterokont' er synonymer. De er ikke fordi de er definert annerledes og til tross for sammensetningsoverlapping, innebærer de fleste anvendelser av navnene forskjellige sammensetninger.
Morfologi
Motile celler

Mange heterokoner er encellede flagellater , og de fleste andre produserer flagellerte celler på et tidspunkt i livssyklusen, for eksempel som gameter eller zoosporer . Navnet heterokont refererer nå til den karakteristiske formen for disse cellene, som vanligvis har to ulik flageller. Den fremre flagellum er dekket med en eller to rader med laterale hår eller mastigonemer , som er trepartige (dvs. med en fleksibel basal innsetting, en stiv hul komponent og tippet med fine fine hår), mens den bakre flagellum er glatt og vanligvis kortere , eller noen ganger redusert til en basal kropp. Flagellene settes inn subapisk eller lateralt, og støttes vanligvis av fire mikrotubuli -røtter i et særegent mønster. Opaliner har mange rader med flagella som ikke har flagellhår. De er nært beslektet med endosymbiotiske proteromonad flagellater hvorav noen har trepartshår som strekker seg fra kroppsoverflaten.
Mastigonemer produseres av glykoproteiner i endoplasmatisk retikulum før de transporteres til den fremre flagellære overflaten. Når hårete flagellum slår, blir de stive mastigonemene tvunget bakover, og dette skaper en retrograd strøm, trekker cellen gjennom vannet eller bringer inn mat.
Begrepet mastigonemer refererer til forskjellige typer flagellhår, men de til stramenopiler har en særegen struktur. Den ble behandlet som den evolusjonære innovasjonen som definerte stramenopilene, og selv om den ikke var det i utgangspunktet, blir den i økende grad behandlet som den definerende egenskapen til heterokontene. Mastigonemer har gått tapt i noen få heterkontlinjer, særlig diatomer, opaliner og actinphryid heliozoa.
Kloroplaster
Mange heterokoner er alger med kloroplaster omgitt av fire membraner, som telles fra den ytterste til den innerste membranen. Den første membranen er kontinuerlig med vertens kloroplast endoplasmatiske retikulum , eller cER. Den andre membranen presenterer en barriere mellom lumen i cER og den primære endosymbiont eller kloroplast, som representerer de to neste membranene, i hvilke thylakoidmembranene finnes. Denne ordningen av membraner har ført til hypotesen om at heterokont kloroplaster ble innhentet fra reduksjon av et symbiotisk rød alge eukaryote, som hadde oppstått ved evolusjonære divergens fra monophyletic primære endosymbiotic stamfar som er antatt å ha gitt opphav til alle eukaryote photoautotrophs . Kloroplastene inneholder karakteristisk klorofyll a og klorofyll c , og vanligvis tilleggspigmentet fucoxanthin , noe som gir dem en gyldenbrun eller brungrønn farge. På grunn av denne fargen blir de referert til som 'kromoplaster' som skiller dem fra klorofyll B som inneholder plastider av grønne alger, deres etterkommere de høyere plantene og euglenider.
De fleste basale heterokontene er fargeløse. Dette antyder at de divergerte før oppkjøpet av kloroplaster i gruppen. Imidlertid finnes fukoksantinholdige kloroplaster også blant haptofytene . Dette førte til hypoteser om at alle organismer med klorofyll a/c som inneholder kloroplaster har en felles fylogenetisk historie med kryptomonader , og bør grupperes taksonomisk som Chromista . Molekylære studier bekrefter ikke at stramenopilene, haptofytene og kryptomonadene er søstertaxa. Den nåværende konsensus er at de forfedre stramenopilene / heterokontene var heterotrofe og skaffet seg kloroplaster etter at deres definerende trekk (trepartshårene) dukket opp.
Klassifisering
Som nevnt ovenfor varierer klassifiseringen betraktelig. Opprinnelig ble heterokontalgen bare brukt for Xanthophytes. Konseptet omformet til å omfatte flere slektninger og ble av noen ansett som en del av kongeriket Plantae og senere, av andre, som i Protista. Et eksempel er:
- Divisjon Chrysophyta
- Divisjon Phaeophyta ( brunalger )
I denne ordningen er Chrysophyceae unnfanget av en veldig omfattende gruppe (Chrysophyte sensu lato ) som var parafyletisk - ettersom diatomer og brunalger utviklet seg fra krysofytene. Over tid har forskjellige avstamninger fått sine egne klasser og ofte divisjoner. Nylige systemer behandler disse ofte som klasser innenfor en enkelt divisjon, kalt Heterokontophyta, Chromophyta eller Ochrophyta. Dette er imidlertid ikke universelt; Round et al. behandle kiselalger som en inndeling.
Oppdagelsen av at oomycetes og hyphochytrids er relatert til disse algene, i stedet for sopp, som tidligere antatt, har ført til at mange forfattere har inkludert disse to gruppene blant heterokontene. Skulle det vise seg at de utviklet seg fra fargede forfedre, ville heterokont -gruppen være parafyletisk i deres fravær. Nok en gang varierer bruken. David J. Patterson kalte denne utvidede gruppen stramenopilene, preget av tilstedeværelsen av trepartsmastigonemer, mitokondrier med rørformede cristae og åpen mitose . Han brukte stramenopilene som en prototype for en klassifisering uten Linnean rang. Sammensetningen har i hovedsak vært stabil, men bruken innenfor rangerte systemer varierer.
Thomas Cavalier-Smith behandler heterokontene som identiske i sammensetningen med stramenopilene; dette er definisjonen som følges her. Han har foreslått å plassere dem i et eget rike, Chromalveolata , sammen med haptofytter, kryptomonader og alveolater. Dette er en av de vanligste revisjonene av systemet med fem riker , men har ikke blitt vedtatt, fordi Chromalveolata ikke er en monofyletisk gruppe. Noen få behandler Chromalveolata som identiske i sammensetningen med heterokontene, eller angir dem som et rike Stramenopila.
Noen kilder deler heterokontene inn i den autotrofiske Ochrophyta og heterotrofiske Bigyra og Pseudofungi . Noen endringer i disse klassifiseringene har imidlertid blitt foreslått.
Navnet Heterokonta kan forveksles med (mye eldre) navnet Heterokontae, som generelt tilsvarer Xanthophyceae , en begrenset delmengde av Heterokonta.
Den forenklede klassifiseringen av gruppen ifølge Adl et al. (2012), der heterokoner (ved å bruke begrepet Stramenopiles) er en del av en større klade som omfatter Alveolates og Rhizaria , er:
-
SAR
-
Stramenopiles
Patterson , 1989, emend. Adl et al. , 2005
- Opalinata Wenyon , 1926, EMEND. Cavalier-Smith , 1997 ( Slopalinida Patterson , 1985)
- Blastocystis Alexeev , 1911
- Bicosoecida Grassé , 1926, emend. Karpov , 1998
- Placidida Moriya et al. , 2002
- Labyrinthulomycetes Dick , 2001
- Hyphochytriales Sparrow , 1960
- Peronosporomycetes Dick , 2001 ( Öomycetes Winter , 1897, emend. Dick , 1976)
- Actinophryidae Claus 1874, emend. Hartmann 1926
- Bolidomonas Guillou & Chrétiennot-Dinet , 1999 ( Bolidophyceae i Guillou et al., 1999)
- Chrysophyceae Pascher , 1914
- Dictyochophyceae Silva , 1980
- Eustigmatales Hibberd , 1981
- Olisthodiscophyceae Barcytė, Eikrem & M.Eliáš , 2021
- Pelagophyceae Andersen & Saunders , 1993
- Phaeothamniophyceae Andersen & Bailey , i Bailey et al. , 1998
- Pinguiochrysidales Kawachi et al. , 2003
- Raphidophyceae Chadefaud , 1950, emend. Silva , 1980
- Synurales Andersen , 1987
- Xanthophyceae Allorge , 1930, emend. Fritsch , 1935 ( Heterokontae Luther , 1899, Heteromonadea Leedale , 1983, Xanthophyta Hibberd , 1990)
- Phaeophyceae Hansgirg , 1886 (ikke Kjellman , 1891, ikke Pfitzer , 1894)
- Schizocladia Henry et al. , i Kawai et al. , 2003 ( Schizocladales Kawai et al. , 2003) (M)
- Diatomea Dumortier , 1821 (= Bacillariophyta Haeckel , 1878)
- Alveolata
- Rhizaria
-
Stramenopiles
Patterson , 1989, emend. Adl et al. , 2005
Filogeni
Basert på følgende verk av Ruggiero et al. 2015 og Silar 2016.
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Galleri
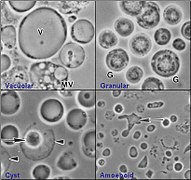
Blastocystis hominis (Blastocystea)

Poteter med Phytophthora ( Oomycetes )

Actinophrys sol ( Actinophryida )

Nannochloropsis sp. ( Eustigmatophyceae )

Dinobryon sp. ( Chrysophyceae )

Synura sp. ( Synurophyceae )

Vaucheria ( Xanthophyta )

Pelagophycus porra ( Phaeophyceae )