
Brodmann -området - Brodmann area
| Brodmann -området | |
|---|---|
 3D -representasjon av Brodmann -områder
| |
| Detaljer | |
| Del av | Cerebrum |
| Identifikatorer | |
| NeuroNames | 427 |
| FMA | 68596 |
| Anatomiske termer for neuroanatomi | |
En Brodmann område er et område av hjernebarken , i den menneskelige eller annen primat hjerne , definert ved sin cytoarchitecture , eller histologisk struktur og organisering av celler .
Historie

Brodmann -områder ble opprinnelig definert og nummerert av den tyske anatom Korbinian Brodmann basert på den cytoarchitectural organisasjonen av nevroner han observerte i hjernebarken ved bruk av Nissl -metoden for cellefarging. Brodmann publiserte sine kart over kortikale områder i mennesker, aper og andre arter i 1909, sammen med mange andre funn og observasjoner om de generelle celletyper og laminær organisering av pattedyr cortex . Det samme Brodmann -områdetallet i forskjellige arter indikerer ikke nødvendigvis homologe områder. Et lignende, men mer detaljert kortisk kart ble utgitt av Constantin von Economo og Georg N. Koskinas i 1925.
Nåværende betydning
Brodmann -områder har blitt diskutert, debattert, foredlet og omdøpt uttømmende i nesten et århundre og forblir den mest kjente og ofte siterte cytoarchitectural organisasjonen av den menneskelige cortex.
Mange av områdene Brodmann definerte utelukkende basert på deres nevronale organisasjon, har siden blitt korrelert tett med forskjellige kortikale funksjoner. For eksempel er Brodmann -områdene 3, 1 og 2 den primære somatosensoriske cortex ; område 4 er den primære motoriske cortex ; område 17 er den primære visuelle cortex ; og områdene 41 og 42 korresponderer tett med primær auditiv cortex . Høyere ordenfunksjoner i assosiasjonens kortikale områder er også konsekvent lokalisert til de samme Brodmann -områdene ved nevrofysiologisk , funksjonell avbildning og andre metoder (f.eks. Den konsekvente lokaliseringen av Brocas tale- og språkområde til venstre Brodmann -områdene 44 og 45 ). Imidlertid kan funksjonell avbildning bare identifisere den omtrentlige lokaliseringen av hjerneaktiveringer når det gjelder Brodmann -områder siden deres faktiske grenser i enhver individuell hjerne krever dens histologiske undersøkelse.
Oversikt

Ulike deler av hjernebarken er involvert i forskjellige kognitive og atferdsfunksjoner. Forskjellene viser seg på en rekke måter: effekten av lokalisert hjerneskade, regionale aktivitetsmønstre avslørt når hjernen undersøkes ved bruk av funksjonelle bildeteknikker, tilkobling med subkortikale områder og regionale forskjeller i cortexens cellulære arkitektur. Nevrovitere beskriver det meste av cortex - delen de kaller neocortex - som har seks lag, men ikke alle lag er synlige i alle områder, og selv når et lag er tilstede, kan dets tykkelse og mobilorganisasjon variere. Forskere har konstruert kart over kortikale områder på grunnlag av variasjoner i lagers utseende sett med et mikroskop. En av de mest brukte ordningene kom fra Korbinian Brodmann , som delte cortex i 52 forskjellige områder og tildelte hver et nummer (mange av disse Brodmann -områdene har siden blitt delt inn). For eksempel er Brodmann -område 1 den primære somatosensoriske cortex, Brodmann -område 17 er den primære visuelle cortex, og Brodmann -område 25 er den fremre cingulære cortex.

Mange av disse hjerneområdene definert av Brodmann har sine egne komplekse interne strukturer. I en rekke tilfeller er hjerneområder organisert i topografiske kart , hvor tilstøtende biter av cortex tilsvarer tilgrensende deler av kroppen, eller av en mer abstrakt enhet. Et enkelt eksempel på denne typen korrespondanse er den primære motoriske cortex, en vevstrimmel som løper langs den fremre kanten av den sentrale sulcus. Motorområder som innerverer hver del av kroppen stammer fra en distinkt sone, med nabokroppsdeler representert av nabosoner. Elektrisk stimulering av cortex når som helst forårsaker muskelsammentrekning i den representerte kroppsdelen. Denne "somatotopiske" representasjonen er imidlertid ikke jevnt fordelt. Hodet, for eksempel, er representert av et område som er omtrent tre ganger så stort som sonen for hele ryggen og stammen. Størrelsen på en hvilken som helst sone korrelerer med presisjonen til motorstyring og mulig sensorisk diskriminering. Områdene for lepper, fingre og tunge er spesielt store, med tanke på proporsjonal størrelse på de representerte kroppsdelene.
I visuelle områder er kartene retinotopiske ; dette betyr at de gjenspeiler topografien på netthinnen , laget av lysaktiverte nevroner langs øyets bakside. Også i dette tilfellet er representasjonen ujevn: fovea - området i midten av synsfeltet - er sterkt overrepresentert i forhold til periferien. Den visuelle kretsen i den menneskelige hjernebarken inneholder flere titalls forskjellige retinotopiske kart, som hver er viet til å analysere den visuelle inngangsstrømmen på en bestemt måte. Den primære visuelle cortex (Brodmann -område 17), som er hovedmottakeren for direkte innspill fra den visuelle delen av thalamus, inneholder mange nevroner som lettest aktiveres av kanter med en bestemt orientering som beveger seg over et bestemt punkt i synsfeltet. Visuelle områder lenger nedstrøms trekker ut funksjoner som farge, bevegelse og form.
I hørselsområder er det primære kartet tonotopisk . Lyder analyseres i henhold til frekvens (dvs. høy tonehøyde vs. lav tonehøyde) av subkortiske hørselsområder, og denne analysen gjenspeiles av den primære hørselssonen i cortex. Som med det visuelle systemet, er det en rekke tonotopiske kortikale kart, som hver er viet til å analysere lyd på en bestemt måte.
Innenfor et topografisk kart kan det noen ganger være finere nivåer av romlig struktur. I den primære visuelle cortex, for eksempel, hvor hovedorganisasjonen er retinotopisk og hovedresponsene er på bevegelige kanter, blir celler som reagerer på forskjellige kantorienteringer, romlig skilt fra hverandre.
For mennesker og andre primater
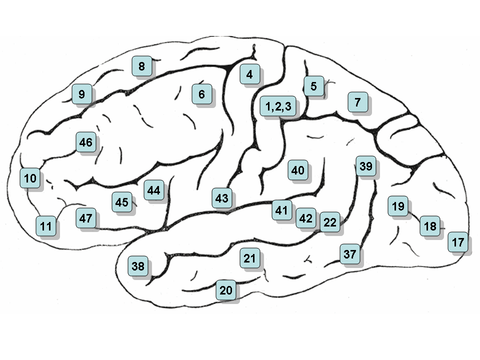
- Område 3, 1 og 2 - Primær somatosensorisk cortex i den postcentrale gyrus (ofte referert til som områder 3, 1, 2 etter konvensjon)
- Område 4 - Primær motorisk cortex
- Område 5 - Superior parietal lobule
- Område 6 - Premotorisk cortex og Supplementary Motor Cortex (Secondary Motor Cortex) ( Supplerende motorområde )
- Område 7-Visuo-Motorkoordinering
- Område 8 - Inkluderer frontale øyefelt
- Område 9 - Dorsolateral prefrontal cortex
- Område 10 - Anterior prefrontal cortex (mest rostral del av overlegen og midtre frontal gyri)
- Område 11 - Orbitofrontal område (orbital og rectus gyri, pluss en del av den rostrale delen av den overlegne frontale gyrus)
- Område 12 - Orbitofrontal område (pleide å være en del av BA11, refererer til området mellom den overlegne frontale gyrus og den dårligere rostrale sulcus)
- Område 13 og område 14 * - Insular cortex
- Område 15 * - Anterior Temporal lobe
- Område 16 - Insular cortex
- Område 17 - Primær visuell cortex (V1)
- Område 18 - Sekundær visuell cortex (V2)
- Område 19 - Associativ visuell cortex (V3, V4, V5)
- Område 20 - Inferior temporal gyrus
- Område 21 - Midt -timelig gyrus
- Område 22 - En del av den overordnede temporale gyrus , inkludert i Wernickes område
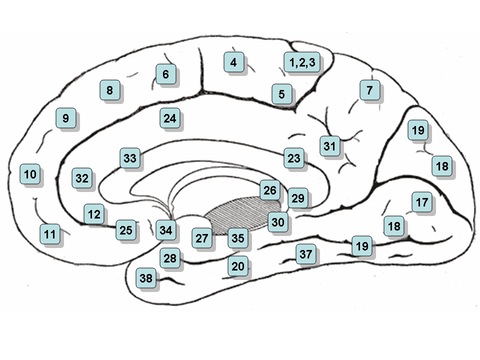
- Område 23 - Ventral posterior cingulate cortex
- Område 24 - Ventral fremre cingulat cortex .
- Område 25 - Subgenuelt område (del av ventromedial prefrontal cortex )
- Område 26 - Ektosplenial del av retrosplenialområdet i hjernebarken
- Område 27 - Presubiculum
- Område 28 - Ventral entorhinal cortex
- Område 29 - Retrosplenial cortex
- Område 30 - Underavdeling av retrosplenial cortex
- Område 31 - Dorsal posterior cingulate cortex
- Område 32 - Dorsal fremre cingulat cortex
- Område 33 - Del av fremre cingulat cortex
- Område 34 - Dorsal entorhinal cortex (på Parahippocampal gyrus )
- Område 35 - Del av perirhinal cortex (i rhinal sulcus )
- Område 36 - Del av perirhinal cortex (i rhinal sulcus )
- Område 37 - Fusiform gyrus
- Område 38 - Temporopolar område (mest rostral del av superior og midterste temporal gyri)
- Område 39 - Vinkelgyrus , av noen ansett for å være en del av Wernickes område
- Område 40 - Supramarginal gyrus som av noen anses å være en del av Wernickes område
- Områder 41 og 42 - Auditiv cortex
- Område 43 - Primær gustatorisk cortex
- Områder 44 og 45 - Brocas område , inkluderer operkulær del og trekantet del av nedre frontal gyrus
- Område 46 - Dorsolateral prefrontal cortex
- Område 47 - Orbital del av dårligere frontal gyrus
- Område 48 - Retrosubikulært område (en liten del av den mediale overflaten av tinninglappen)
- Område 49 - Parasubikulært område i en gnager
- Område 52 - Parainsular område (i krysset mellom tinninglappen og insulaen )
(*) Område bare funnet i ikke-menneskelige primater .
Noen av de opprinnelige Brodmann -områdene har blitt delt videre, f.eks. "23a" og "23b".
Klikkbart kart: sideflate
- Merk: sidevisningen eller sidevisningen av hjernen er betegnet 'sideflaten'

Klikkbart kart: medial overflate
- Merk: utsikten over snittet mellom høyre og venstre hjernehalvdel i hjernen er betegnet 'medial overflate'

Kritikk
Da von Bonin og Bailey konstruerte et hjernekart for makak -apen, fant de beskrivelsen av Brodmann utilstrekkelig og skrev: "Brodmann (1907), det er sant, utarbeidet et kart over den menneskelige hjerne som har blitt gjengitt mye, men dessverre, dataene de ble basert på ble aldri publisert "De brukte i stedet cytoarchitechtonic -opplegget til Constantin von Economo og Georg N. Koskinas publisert i 1925 som hadde den" eneste akseptable detaljerte beskrivelsen av den menneskelige cortex ".
Se også
Referanser
Eksterne linker
- [1] - Brodmann -områder, deres funksjoner og lateralisering av funksjoner på tvers av halvkule
- Brodmann , Mark Dubin sider om Brodmann -områder.
- Brodmann -områder Brodmann -områder av cortex involvert i språk.
- Brodmann Illustrations BrainInfo Illustrations.