
CD9 - CD9
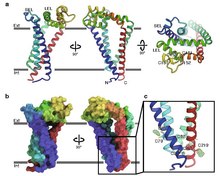





CD9 er et gen som koder for et protein som er medlem av transmembrane 4 -superfamilien også kjent som tetraspanin -familien. Det er en glykoprotein på celleoverflaten som består av fire transmembrane regioner og har to ekstracellulære sløyfer som inneholder disulfidbindinger som er bevart i hele tetraspanin -familien. Inneholder også forskjellige palmitoyleringssteder som lar CD9 samhandle med lipider og andre proteiner.
Funksjon
Tetraspaninproteiner er involvert i en rekke biologiske prosesser som vedheft, motilitet, membransmelting, signalering og proteinhandel. Tetraspaniner spiller en rolle i mange biologiske prosesser på grunn av deres evne til å samhandle med mange forskjellige proteiner, inkludert interaksjoner mellom hverandre. Deres distinkte palmitoyleringssteder lar dem organisere seg på membranen til tetraspaninberikede mikrodomener (TEM). Disse TEMene antas å spille en rolle i mange cellulære prosesser, inkludert exosome biogenese. CD9 brukes ofte som en markør for eksosomer ettersom det er på overflaten.
Imidlertid spiller CD9 i noen tilfeller en større rolle i eksosomers evne til å være mer eller mindre patogen. Eksosomer er vist i HIV-1-infeksjon og er i stand til å forbedre HIV-1-inntreden gjennom tetraspanin CD9 og CD81. Imidlertid ser det ut til at uttrykk for CD9 på cellemembranen reduserer viral inntreden av HIV-1.
CD9 har en mangfoldig rolle i cellulære prosesser, da det også har vist seg å utløse blodplateaktivering og aggregering. Det danner et alphaIIbbeta3-CD9-CD63-kompleks på overflaten av blodplater som interagerer direkte med andre celler, for eksempel nøytrofiler som kan hjelpe i immunrespons. I tillegg ser det ut til at proteinet fremmer fusjon av muskelceller og støtter vedlikehold av myotube. Spiller også en nøkkelrolle i egg-sædfusjon under befruktning av pattedyr. Mens oocytter har eggløsning, smelter ikke oocytter med CD9-mangel på riktig måte med sæd ved befruktning. CD9 er lokalisert i mikrovillarmembranen i oocyttene og ser også ut til å gripe inn for å opprettholde normal form av oocyttmikrovilli .
CD9 kan også modulere celleadhesjon og migrasjon. Denne funksjonen gjør CD9 interessant når man studerer kreft og kreftmetastaser. Imidlertid ser det ut til at CD9 har en varierende rolle i ulike typer kreft. Studier viste at CD9 -ekspresjonsnivåer har en omvendt korrelasjon til metastatisk potensial eller pasientoverlevelse. Overuttrykket av CD9 ble vist å redusere metastaser i visse typer melanom, bryst, lunge, bukspyttkjertel og tykktarmskreft. Men i andre studier har CD9 vist seg å øke migrasjonen eller komme sterkt til uttrykk i metastatisk kreft i forskjellige cellelinjer som lungekreft, magekreft av scirrhous type, hepatocellulært karsinom, akutt lymfoblastisk leukemi og brystkreft. Forslag basert på kreft CD9 kan være en svulstundertrykkende eller promotor. Det har også blitt antydet at CD9 har en effekt på kreftcelles evne til å utvikle kjemoresistens.
I tillegg har CD9 vist seg å blokkere vedheft av Staphylococcus aureus til sår. Vedheftet er avgjørende for infeksjon av såret. Dette antyder at CD9 kan være nyttig som behandling for hudinfeksjon av Staphylococcus aureus.
Interaksjoner
CD9 har vist seg å samhandle med:
Se også
Referanser
Videre lesning
- Horejsí V, Vlcek C (august 1991). "Ny strukturelt distinkt familie av leukocyttoverflateglykoproteiner inkludert CD9, CD37, CD53 og CD63" . FEBS Letters . 288 (1–2): 1–4. doi : 10.1016/0014-5793 (91) 80988-F . PMID 1879540 . S2CID 26316623 .
- Berditchevski F (desember 2001). "Komplekser av tetraspaniner med integriner: mer enn man ser for øyet". Journal of Cell Science . 114 (Pt 23): 4143–51. doi : 10.1242/jcs.114.23.4143 . PMID 11739647 .
- Ninomiya H, Sims PJ (juli 1992). "Det humane komplement-regulatoriske proteinet CD59 binder seg til alfa-kjeden til C8 og til" b "-domenet til C9" . Journal of Biological Chemistry . 267 (19): 13675–80. doi : 10.1016/S0021-9258 (18) 42266-1 . PMID 1377690 .
- Miyake M, Koyama M, Seno M, Ikeyama S (desember 1991). "Identifikasjon av det motilitetsrelaterte proteinet (MRP-1), gjenkjent av monoklonalt antistoff M31-15, som hemmer cellemotilitet" . Journal of Experimental Medicine . 174 (6): 1347–54. doi : 10.1084/jem.174.6.1347 . PMC 2119050 . PMID 1720807 .
- Boucheix C, Benoit P, Frachet P, Billard M, Worthington RE, Gagnon J, Uzan G (januar 1991). "Molekylær kloning av CD9 -antigenet. En ny familie av celleoverflateproteiner" . Journal of Biological Chemistry . 266 (1): 117–22. doi : 10.1016/S0021-9258 (18) 52410-8 . PMID 1840589 .
- Iwamoto R, Senoh H, Okada Y, Uchida T, Mekada E (oktober 1991). "Et antistoff som hemmer bindingen av difteritoksin til celler avslørte forbindelsen mellom et 27 kDa membranprotein og difteritoksinreseptoren" . Journal of Biological Chemistry . 266 (30): 20463–9. doi : 10.1016/S0021-9258 (18) 54947-4 . PMID 1939101 .
- Benoit P, Gross MS, Frachet P, Frézal J, Uzan G, Boucheix C, Nguyen VC (januar 1991). "Tildeling av det humane CD9 -genet til kromosom 12 (region P13) ved bruk av humane spesifikke DNA -prober". Menneskelig genetikk . 86 (3): 268–72. doi : 10.1007/bf00202407 . PMID 1997380 . S2CID 27178985 .
- Lanza F, Wolf D, Fox CF, Kieffer N, Seyer JM, Fried VA, et al. (Juni 1991). "cDNA-kloning og ekspresjon av blodplate p24/CD9. Bevis for en ny familie av flere membranoverspennende proteiner" . Journal of Biological Chemistry . 266 (16): 10638–45. doi : 10.1016/S0021-9258 (18) 99271-9 . PMID 2037603 .
- Higashihara M, Takahata K, Yatomi Y, Nakahara K, Kurokawa K (mai 1990). "Rensing og delvis karakterisering av CD9 -antigen av humane blodplater". FEBS Letters . 264 (2): 270–4. doi : 10.1016/0014-5793 (90) 80265-K . PMID 2358073 . S2CID 42129059 .
- Masellis-Smith A, Shaw AR (mars 1994). "CD9-regulert vedheft. Monoklonalt anti-CD9-antistoff induserer pre-B-celleadhesjon til beinmargsfibroblaster gjennom de novo-gjenkjenning av fibronektin". Journal of Immunology . 152 (6): 2768–77. PMID 7511626 .
- Chalupny NJ, Kanner SB, Schieven GL, Wee SF, Gilliland LK, Aruffo A, Ledbetter JA (juli 1993). "Tyrosinfosforylering av CD19 i pre-B og modne B-celler" . EMBO Journal . 12 (7): 2691–6. doi : 10.1002/j.1460-2075.1993.tb05930.x . PMC 413517 . PMID 7687539 .
- Rubinstein E, Benoit P, Billard M, Plaisance S, Prenant M, Uzan G, Boucheix C (april 1993). "Organisering av det humane CD9 -genet". Genomikk . 16 (1): 132–8. doi : 10.1006/geno.1993.1150 . PMID 8486348 .
- Schmidt C, Künemund V, Wintergerst ES, Schmitz B, Schachner M (januar 1996). "CD9 av musens hjerne er involvert i neurittvekst og cellemigrasjon in vitro og er assosiert med alfa 6/beta 1 -integrinet og det nevrale vedheftsmolekylet L1". Journal of Neuroscience Research . 43 (1): 12–31. doi : 10.1002/jnr.490430103 . PMID 8838570 . S2CID 84774340 .
- Sincock PM, Mayrhofer G, Ashman LK (april 1997). "Lokalisering av transmembrane 4 superfamilie (TM4SF) medlem PETA-3 (CD151) i normale humane vev: sammenligning med CD9, CD63 og alfa5beta1 integrin" . Journal of Histochemistry and Cytochemistry . 45 (4): 515–25. doi : 10.1177/002215549704500404 . PMID 9111230 .
- Rubinstein E, Poindessous-Jazat V, Le Naour F, Billard M, Boucheix C (august 1997). "CD9, men ikke andre tetraspaner, assosierer seg med beta1 -integrinforløperen". European Journal of Immunology . 27 (8): 1919–27. doi : 10.1002/eji.1830270815 . PMID 9295027 . S2CID 42866423 .
- Cho, JH, Kim, E., Son, Y. et al. (2020). CD9 induserer cellulær aldring og forverrer aterosklerotisk plakkdannelse. Celledød og differensiering https://doi.org/10.1038/s41418-020-0537-9
Eksterne linker
- Menneskelig CD9 genomlokalisering og CD9 -gen detaljer side i UCSC Genome Browser .