
Proteaceae - Proteaceae
| Proteaceae | |
|---|---|

|
|
| Blomstring av Protea cynaroides | |
|
Vitenskapelig klassifisering |
|
| Kongedømme: | Plantae |
| Clade : | Trakeofytter |
| Clade : | Angiospermer |
| Clade : | Eudikoter |
| Rekkefølge: | Proteales |
| Familie: |
Proteaceae Juss. |
| Slekter | |
|
Ca 80, se tekst |
|
Den Proteaceae / ˌ p r oʊ t i eɪ s jeg / danner en familie av blomsterplanter hovedsakelig fordelt i den sørlige halvkule . Familien består av 83 slekter med ca 1660 kjente arter . Sammen med Platanaceae og Nelumbonaceae utgjør de ordenen Proteales . Kjente slekter inkluderer Protea , Banksia , Embothrium , Grevillea , Hakea og Macadamia . Arter som New South Wales waratah ( Telopea speciosissima ), king protea ( Protea cynaroides ) og forskjellige arter av Banksia , soman og Leucadendron er populære snittblomster . Nøttene til Macadamia integrifolia vokser mye kommersielt og konsumeres, i likhet med de av Gevuina avellana i mindre skala. Australia og Sør -Afrika har de største konsentrasjonene av mangfold.
Etymologi
Navnet Proteaceae av Antoine Laurent de Jussieu i 1789 var basert på slekten Protea , som i 1767 Carl Linnaeus stammer fra navnet på den greske guden Proteus , en guddom som var i stand til å skifte mellom mange former. Dette er et passende bilde, ettersom familien er kjent for sin forbløffende variasjon og mangfold av blomster og blader.
Beskrivelse

Slektene til Proteaceae er svært varierte, med Banksia spesielt et slående eksempel på adaptiv stråling i planter. Denne variasjonen gjør det umulig å gi en enkel, diagnostisk identifikasjonsnøkkel for familien, selv om individuelle slekter lett kan identifiseres.
- Proteaceae området fra nedbrutt busker til høye skogstrær , av 40 m i høyde, og er vanligvis middels høy eller lav eller flerårige busker , med unntak av noen Stirlingia arter som er urter . Noen arter er fakultativt løvfellende ( Embothrium coccineum ), sjelden acaulescent, cauline -delen av kragen blir ofte tykkere ( lignotuber ). Indumentum av trecellede hår, noen ganger kjertel, sjelden fraværende, den apikale cellen er vanligvis langstrakt, akutt, noen ganger like eller ulikt bifid.
- Blader sjelden aromatiske, vanligvis vekslende, og i en spiral, sjelden motsatt eller vertikal; coriaceous, sjelden kjøttfull eller spinescent, enkel eller sammensatt (imparipinate, imparibipinate eller sjelden palmate eller digitate med pinnatisect segmenter), hele kanten til (3-) pinnatisect (gir en bregner-lignende aspekt); sjelden delt todelt, ofte fjerntannet, krenet eller takket, sittende eller stilket; den petiole ofte med en hoven base, men sjelden omsluttet (noen ganger i Synaphea ), uten stipules ; finnete og til håndformet eller parallell venation , brochidodromous eller redusert til et enkelt fremtredende skovl, vernation normalt conduplicate; anisofyl forekommer ofte i de forskjellige vekstperioder; bladblad dorsiventral, isobilateral eller sentrert; mesofyllvev vanligvis med sclerenchymatøse idioblaster , sjeldne sekretoriske hulrom. Brachy-paracytic stomata (laterocytisk i Bellendena ).
Plantestammene med to typer av radier, brede og multi-taggete eller smale og uni-oppriflet barken stratifisert eller ikke, trilacunar noder med tre blad spor (sjelden unilacunar med en trase), sclereids hyppig; bark med lenticeller ofte horisontalt forstørret, korkkambium til stede, vanligvis overfladisk. Røtter lateralt og kort, ofte gruppert i bunter ( proteoidrøtter ) med veldig tette rothår , sjelden med mycorrhiza .
- Planter er vanligvis hermafrodittiske, mer sjelden monoecious, dioecious eller andromonoecious.
- Blomsterstander meget variabel, enkle eller sammensatte, aksillære eller terminal, laterale blomster enslig eller i par, sjelden med en terminal blomst, racemiform , paniculate eller kondensert, vanligvis med dekkbladene , noen ganger omdannes til blader eller squamiform, som danner en slags membran, eller med lyse farger, som danner en involucre eller pseudanthium, peduncles og pedicels noen ganger kontrakt, komprimert med rachis , i noen tilfeller de overbelastede blomsterstanden danner super blomsterstander (noen Alloxylon ); svært sjelden er blomstene ensomme og aksillære nær enden av grener; hos arter med lignotubers vokser blomstene noen ganger fra disse og passerer gjennom jorden (geofytter).
- Blomster er vanligvis perfekte, aktinomorfe eller zygomorfe, hypogynøse, ofte store og prangende. Flat eller skrå, noen ganger danner en gynofor. Hypogyn disk tilstede og ekstrastaminal eller fraværende. Perianth av (3-) 4 (-8) tepals (noen ganger tolket som en dimer og diklamyd perianth), i 1 (-2) ventilerende hvirvler, noen ganger forlenget i en basalsekk, fri eller smeltet på forskjellige måter (alle smeltet eller til og med en ledig og tre basalt til helt sammensmeltet), eller til og med konnivent ved marginalt interdigiterte papiller som danner et rør eller en bilabiat struktur, zygomorfe, noen ganger åpnes lateralt på en rekke måter. Haplostemonous androecium, vanligvis isostemonous, motsatt av (3-) 4 (-5) støvdragere , alle fruktbare eller noen omdannet til staminoder , vanligvis filamentøse, filamenter delvis eller helt smeltet til tepalene, sjelden gratis, basifiksede anthers adnate, ditheous, tetrasporangiate, noen ganger unilokulær og bisporagiat, intror til latrorse (sjelden), utvidet bindebånd, vanligvis med apiculus, dehiscence langs langsgående tårer. Hypogyniske kjertler (0-) 1-4, squamiform eller langstrakte, kjøttfulle, frie eller smeltede og danner en gal eller ringformet nektar over beholderen. Overlegen gynoecium av 1 (-2) apocarpous carpels , fastsittende eller stipitate (med en mer eller mindre langstrakt gynophore ), noen ganger ikke er fullstendig lukket, stil vanligvis utvikles, stigma liten eller i form av en klemme eller terminal under disk eller til og med sideveis og skrå, ofte innrykket, papiløs, fuktig eller tørr, eggløsninger 1-100 eller mer per karpel, anatropøs, hemianatropøs, amfitropøs eller ortotropøs, for det meste hemitropøs, bitegmisk, crassinucellat, chalaza med en ring av vaskulære bunter, funiculus er tidvis fraværende og ovulum er kondensert til placenta, marginal placentation med forskjellige disposisjoner eller apikale.
- Frukt dehiscent eller indehiscent, i achene eller kjerne, follicle , drupe (med lignified endocarp) eller falskt drupal (med lignified indre mesocarp), noen ganger lik en caryopsis som det er smeltet til eggstokkens vegg og testa, ofte lignified og serotinøs; frukten fra den samme blomsterstanden er noen ganger smeltet og danner en synkarp.
- Frø 1-mange, noen ganger vingede, flate til avrundede, med endosperm fraværende, tilstede i Bellendina , endotesta med et uvanlig lag som inneholder krystaller av kalsiumoksalat som sjelden er fraværende, godt differensiert embryo , rett, dikotyledonøst, men ofte med 3 eller flere ( opptil 9) store cotyledons, ofte aurikulerte.
- Pollen i monader, trekantet i polar utsikt, (2-) 3 (-8) -aperturat, vanligvis isopolar og triporat, biporat i Embothrium og stammen Banksieae, kolpoidat i Beauprea , sfærisk i Aulax og Franklandia eller sterkt anisopolar hos noen arter av Persoonia ; åpningene til førstnevnte tetrads følger Garsides lov.
- Kromosomalt tall : n = 5, 7, 10–14, 26, 28; størrelser varierer fra veldig små (gjennomsnitt på 1,0 μm) til veldig store (gjennomsnitt på 14,4 μm) i henhold til art; x = 7, 12.
Blomster

Generelt sett er den diagnostiske egenskapen til Proteaceae det sammensatte blomsterhodet eller, mer nøyaktig, blomsterstand . I mange slekter er den mest åpenbare egenskapen de store og ofte veldig prangende blomsterstandene, bestående av mange små blomster tett pakket inn i et kompakt hode eller pigg. Selv denne karakteren forekommer imidlertid ikke i alle Proteaceae; Adenanthos -arter har for eksempel ensomme blomster. Hos de fleste Proteaceae -arter er pollineringsmekanismen høyt spesialisert. Det innebærer vanligvis bruk av en "pollen-programleder", et område på stil -ende som presenterer pollen til pollinator.
Proteaceae blomsterdeler forekommer i firer, men de fire tepalene er smeltet sammen til et langt, smalt rør med en lukket kopp på toppen, og filamentene til de fire støvdragerne er smeltet til tepalene, på en slik måte at fronter er lukket inne koppen. Støvelen passerer først langs innsiden av perianth -røret, slik at også stigmaet er innelukket i koppen. Etter hvert som blomsten utvikler seg, vokser pistillen raskt. Siden stigmaet er fanget, må stilen bøyes for å forlenges, og til slutt bøyes det så langt, det deler perianthen langs den ene sømmen. Stilen fortsetter å vokse til antesen , når nektarene begynner å produsere nektar . På dette tidspunktet deler perianthen seg i komponentbladene, koppen deler seg, og pistilen slippes ut for å fjære mer eller mindre oppreist.
Økologi

Mange av Proteaceae har spesialiserte proteoidrøtter , masser av laterale røtter og hår som danner en radial absorberende overflate, produsert i bladstrøet under sesongvekst, og vanligvis krymper på slutten av vekstsesongen. De er en tilpasning til vekst i fattige, fosforfattige jordarter, og øker plantens tilgang til mangel på vann og næringsstoffer sterkt ved å utstråle karboksylater som mobiliserer fosfor som tidligere ikke var tilgjengelig. De øker også rotens absorpsjonsoverflate, men dette er et mindre trekk, ettersom det også øker konkurransen om næringsstoffer mot sine egne rotklaser. Men denne tilpasningen later dem svært sårbare for dieback forårsaket av Phytophthora cinnamomi vann mugg , og generelt intolerant av befruktning . På grunn av disse spesialiserte proteoidrøttene, er Proteaceae en av få blomstrende plantefamilier som ikke danner symbioser med arbuskulære mykorrhizale sopp. De utskiller store mengder organiske syrer ( sitronsyre og eplesyre ) hver 2. – 3. Dag for å hjelpe mobilisering og absorpsjon av fosfat. Mange arter er branntilpassede ( pyrofytter ), noe som betyr at de har strategier for å overleve branner som feier gjennom deres habitat. Noen er resproutere , og har en tykk rotstamme begravet i bakken som skyter opp nye stilker etter en brann, og andre er reseeders , noe som betyr at de voksne plantene blir drept av brannen, men sprer frøene sine, som stimuleres av røyken til å ta rot og vokse. Varmen ble tidligere antatt å ha stimulert veksten, men kjemikaliene i røyken har nå vist seg å forårsake den.
Det er fire todommelige slekter ( Aulax , Dilobeia , Heliciopsis og Leucadendron ), 11 andromonoecious slekter og noen andre slekter har arter som er kryptisk andromonoecious: to arter er sterile og reproduserer bare vegetativt ( Lomatia tasmanica , Hakea pulvinifera ). Arten varierer mellom å være autokompatibel og autoinkompatibel, med mellomliggende situasjoner; disse situasjonene forekommer noen ganger i samme art. Blomstene er vanligvis protandrous. Rett før antesen slipper støvknappene pollenet sitt og avsetter det på stigmaet, som i mange tilfeller har et forstørret kjøttfullt område spesielt for avsetning av sitt eget pollen. Nektar-matere kommer neppe til å komme i kontakt med selve stifrene, men kan nesten ikke unngå å ta kontakt med stigmaet; dermed fungerer stigmaet som en pollen-presentatør , og sikrer at nektar-feederne fungerer som pollinatorer. Ulempen med denne pollineringsstrategien er at sannsynligheten for selvbefruktning er sterkt økt; mange Proteaceae motvirker dette med strategier som protandry , selvinkompatibilitet eller foretrukket abort av selfed frø. Systemene for presentasjon av pollen er vanligvis svært forskjellige, tilsvarende diversifiseringen av pollinatorene. Bestøvning utføres av bier , biller , fluer , møll , fugler ( honningspisere , solfugler , sukkerfugler og kolibrier ) og pattedyr (gnagere, små pungdyr , elefantskriver og flaggermus . De to sistnevnte virkemidlene ble evolusjonært avledet fra entomofil i forskjellige, uavhengige hendelser Spredningen av noen arter viser det nysgjerrige fenomenet serotiny , som er forbundet med deres pyrofytiske oppførsel: disse trærne akkumulerer frukt på grenene hvis ytre lag eller beskyttende strukturer ( bladblad ) er svært lignifiserte og motstandsdyktige mot brann. Frukten frigjør bare sine frø når de har blitt brent og når bakken har blitt befruktet med aske fra ilden og er fri for konkurrenter. Mange arter har frø med elaiosomer som er spredt av maur ; frøene med vinger eller tistel ned viser anemokori , mens drupes og andre kjøttfull frukt viser endozoochory når pattedyr og fugler inntar dem. Noen afrikanske og australske gnagere er kjent for å mulatfrukt og frø av disse plantene i reirene for å mate dem, selv om noen klarer å spire.
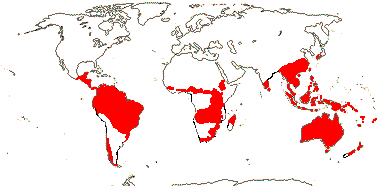
Fordeling
Proteaceae er hovedsakelig en familie på den sørlige halvkule, med sine hovedsenter for mangfold i Australia og Sør -Afrika. Det forekommer også i Sentral-Afrika, Sør- og Mellom-Amerika , India , østlige og sørøstlige Asia og Oseania . Bare to arter er kjent fra New Zealand, selv om bevis på fossilt pollen tyder på at det var flere tidligere.
Det er et godt eksempel på en Gondwanan -familie, med taxa som forekommer på praktisk talt alle landmasser som anses som en rest av det gamle superkontinentet Gondwana, unntatt Antarktis . Det antas at familien og underfamiliene har diversifisert seg godt før fragmenteringen av Gondwana, noe som betyr at alle er godt over 90 millioner år gamle. Bevis for dette inkluderer en overflod av proteaceous pollen funnet i kritt kull forekomster av South Island i New Zealand . Det antas å ha oppnådd sin nåværende distribusjon i stor grad ved kontinentaldrift i stedet for spredning over havgap.
Fytokjemi

Det er ikke utført avgjørende studier på de kjemiske stoffene som finnes i denne brede familien. Slektene Protea og Faurea er uvanlig som de bruker xylose som hoved sukker i sin nektar, og som de har høye konsentrasjoner av polygalactol, mens sukrose er den viktigste sukker tilstede i Grevillea . Cyanogene glykosider , avledet fra tyrosin , er ofte til stede, det samme er proanthocyanidiner ( delphinidin og cyanidin ), flavonoler ( kaempferol , quercetin og myricetin ) og arbutin . Alkaloider er vanligvis fraværende. Iridoider og ellaginsyre er også fraværende. Saponiner og sapogeniner kan enten være til stede eller fraværende hos forskjellige arter. Mange arter akkumulerer aluminium .

Bruksområder og dyrking

Mange tradisjonelle kulturer har brukt Proteaceae som næring, medisin, for å kurere dyrehud, som kilde til fargestoffer, ved og som trevirke for bygging. Aboriginale australiere spiser frukten av Persoonia , og frøene til arter fra andre slekter, inkludert Gevuina og Macadamia , er en del av kostholdet til urbefolkningen, men selges også over hele verden. De ømme skuddene til Helicia -arter brukes på Java, og nektaren fra blomsterstandene til en rekke arter blir drukket i Australia. Tradisjonelle medisiner kan fås fra infusjoner av røtter, bark, blader eller blomster av mange arter som brukes som aktuelle applikasjoner for hudforhold eller internt som tonika, afrodisiakum og galaktogener for å behandle hodepine, hoste, dysenteri, diaré, fordøyelsesbesvær, magesår og nyresykdom. Treet fra trærne i denne familien er mye brukt i konstruksjon og til intern bruk som dekorasjon; treverket fra Protea , Leucadendron og Grevillea er spesielt populært. Mange arter brukes i hagearbeid, spesielt slekter fra Banksia , Embothrium , Grevillea og Telopea . Dessverre har denne bruken resultert i introduksjon av eksotiske arter som har blitt invasive; eksempler inkluderer hakea pil ( Hakea salicifolia ) og silkeaktig hakea ( Hakea sericea ) i Portugal.
To arter av Macadamia dyrkes kommersielt for sine spiselige nøtter. Gevuina avellana (chilensk hassel) dyrkes også for sine spiselige nøtter, i Chile og New Zealand , og de brukes også i farmasøytisk industri for sine fuktighetsbevarende egenskaper og som ingrediens i solkremer . Det er den mest motstandsdyktige mot kulde av trefamiliene som produserer nøtter. Den er også plantet på de britiske øyer og på Stillehavskysten i USA for sitt tropiske utseende og evne til å vokse i kjøligere klima .
Mange arter av Proteaceae dyrkes av barnehageindustrien som barriereplanter og for sine fremtredende og særegne blomster og løvverk. Noen arter er av betydning for snittblomstindustrien , spesielt noen Banksia- og Protea -arter. To arter av slekten Macadamia dyrkes kommersielt for spiselige nøtter.
Sukkerbusk ( Protea ), pincushions ( Leucospermum ) og conebushes ( Leucadendron ), så vel som andre som pagoder ( Mimetes ), Aulax og rødmende bruder ( Serruria ), består av en av de tre viktigste plantegruppene av fynbos , som utgjør en del av Kapp Floral Kingdom , det minste, men rikeste planteriket for sin størrelse og det eneste riket som finnes i et enkelt land. De andre hovedgruppene av planter i fynbos er Ericaceae og Restionaceae . Sør -afrikanske proteas dyrkes dermed mye på grunn av deres mange varierte former og uvanlige blomster. De er populære i Sør -Afrika for sin skjønnhet og sin nytte i dyrelivshager for å tiltrekke seg fugler og nyttige insekter.
Artene som er mest verdsatt som prydplanter er trærne som vokser på sørlige breddegrader da de gir landskap i temperert klima et tropisk utseende; Lomatia ferruginea (Fuinque), Lomatia hirsuta (Rådal) har blitt introdusert i Vest -Europa og den vestlige USA . Embothrium coccineum (Chilean Firetree eller Notro ) er høyt verdsatt på De britiske øyer for sine mørkerøde blomster og kan finnes så langt nord som Færøyene på en breddegrad på 62 ° nord.
Blant banksiaene , hvorav mange vokser i temperert og middelhavsklima, er de aller fleste busker; bare noen få er trær som er verdsatt for høyden. Blant de høyeste artene er: B. integrifolia med dens underarter B. integrifolia subsp. monticola , som er bemerkelsesverdig ettersom plantene som danner underarten er de høyeste trærne i banksiene, og de er mer frostbestandige enn andre banksier, B. seminuda , B. littoralis , B. serrata ; blant de som kan betraktes som små trær eller store busker: B. grandis , B. prionotes , B. marginata , B. coccinea og B. speciosa ; alle disse er plantet i parker og hager og til og med langs veikanter på grunn av deres størrelse. Resten av artene av denne slekten, rundt 170 arter, er busker, selv om noen av dem er verdsatt for blomstene.
En annen art som dyrkes i noen deler av verden, selv om den er mindre, er Telopea speciosissima (Waratah), fra fjellene i New South Wales , Australia .
Noen tempererte klimaarter dyrkes mer lokalt i Australia for sitt attraktive utseende: Persoonia pinifolia ( furublad ) er verdsatt for sine livlige gule blomster og druelignende frukt. Adenanthos sericeus (ullbusk) er plantet for sine attraktive myke blader og sine små røde eller oransje blomster. Hicksbeachia pinnatifolia ( biffnøtt , rød bauple -mutter) plantes ofte for løvverk og spiselige nøtter.
Parasitter

Proteaceae er spesielt utsatt for visse parasitter, spesielt oomycete Phytophthora cinnamomi , som forårsaker alvorlig rotrot i plantene som vokser i middelhavsklima. Fusarium oxysporum forårsaker en sykdom som kalles fusariose i røtter som forårsaker gulning og visning, med alvorlige økologiske skader på skogplanter og økonomiske tap i planter av kommersiell interesse. Andre vanlige infeksjoner er forårsaket av arter av Botryosphaeria , Rhizoctonia , Armillaria , Botrytis , Calonectria og andre sopp.
Bevaringsstatus
Den IUCN vurderer at 47 Proteaceae arter er truet, hvorav én art, Stenocarpus dumbeensis Guillaumin , 1935, fra New Caledonia, er antatt å være utdødd. Arten i denne familien er spesielt utsatt for ødeleggelse eller fragmentering av deres habitat , brann, parasittsykdommer, konkurranse fra introduserte planter, jordforringelse og annen skade forårsaket av mennesker og deres husdyr. Arten påvirkes også av klimaendringer .
Fossiler

Proteaceae har en rik fossilrekord, til tross for de iboende vanskelighetene med å identifisere rester som ikke viser diagnostiske egenskaper. Identifikasjon kommer vanligvis fra bruk av en kombinasjon av brachy-paracytic stomata og de uvanlige trichome- basene eller, i andre tilfeller, den uvanlige strukturen til pollentetrader. Fossiler som kan tilskrives denne familien har blitt funnet på de fleste områdene som dannet Gondwana -superkontinentet. Et bredt utvalg av pollen som tilhører denne familien, dateres tilbake til Upper Cretaceous ( Campanian - Maastrichtian ) fra sørøst i Australia og pollen fra Middle Cretaceous ( Cenomanian - Turonian ) fra Nord -Afrika og Peru beskrevet som Triorites africaensis . De første makrofossilene dukker opp tjue millioner år senere i paleocene i Sør -Amerika og nordøst i Australia. Fossilrekorden for noen områder, for eksempel New Zealand og Tasmania, viser et større biologisk mangfold for Proteaceae enn det som for øyeblikket eksisterer, noe som støtter det faktum at fordelingen av mange taxa har endret seg drastisk med tiden og at familien har lidd en generell nedgang, inkludert høye utryddelsesnivåer under Cenozoic .
Taksonomi

Først beskrevet av den franske botanikeren Antoine Laurent de Jussieu , er familien Proteaceae en ganske stor, med rundt 80 slekter, men mindre enn 2000 arter. Det er anerkjent av praktisk talt alle taksonomer . Fast etablert i henhold til klassiske seksualsystem , er det også anerkjent av cladistics -baserte APG og APG II- systemer. Den er plassert i rekkefølgen Proteales , hvis plassering selv har variert.
En klassifisering av slektene i Proteaceae ble laget av Lawrie Johnson og Barbara Briggs i deres innflytelsesrike monografi fra 1975 " On the Proteaceae: evolusjonen og klassifiseringen av en sørlig familie ", til den i stor grad ble erstattet av molekylære studier av Peter H. Weston og Nigel Barker i 2006. Proteaceae er nå delt inn i fem underfamilier: Bellendenoideae , Persoonioideae , Symphionematoideae , Proteoideae og Grevilleoideae . I 2008 oppdaterte Mast og kolleger Macadamia og relaterte slekter i stammen Macadamieae. Videre ble det funnet at Orites megacarpus ikke var innenfor slekten Orites , og heller ikke i stammen Roupaleae, i stedet i stammen Macadamieae, derav gitt det nye artnavnet Nothorites megacarpus . Hele arrangementet, ifølge Weston og Barker (2006) med oppdateringene til slektene fra Mast et al. (2008), er som følger:
.jpg)
- Familie Proteaceae
- Underfamilien Bellendenoideae
- Underfamilie Persoonioideae
- Stamme Placospermeae
- Stamme Persoonieae
- Underfamilien Symphionematoideae
- Underfamilien Proteoideae
-
incertae sedis
- Eidothea - Beauprea - Beaupreopsis - Dilobeia - Cenarrhenes - Franklandia
- Stamme Conospermeae
- Subtribe Stirlingiinae
- Subtribe Conosperminae
- Stamme Petrophileae
- Stamme Proteeae
- Stamme Leucadendreae
- Subtribbe Isopogoninae
- Subtribe Adenanthinae
- Subtribbe Leucadendrinae
- Leucadendron - Serruria - Paranomus - Vexatorella - Sorocephalus - Spatalla - Leucospermum - Mimetes - Diastella - Orothamnus
-
incertae sedis
- Underfamilien Grevilleoideae
- incertae sedis
- Stamme Roupaleae
-
incertae sedis
- Megahertzia - Knightia - Eucarpha - Triunia
- Subtribe Roupalinae
- Subtribbe Lambertiinae
- Subtribe Heliciinae
- Subtribe Floydiinae
-
incertae sedis
- Stamme Banksieae
- Subtribe Musgraveinae
- Subtribbe Banksiinae
- Stam Embothrieae
- Subtribe Lomatiinae
- Subtribe Embothriinae
- Embothrium - Oreocallis - Alloxylon - Telopea
- Subtribe Stenocarpinae
- Subtribe Hakeinae
- Opisthiolepis - Buckinghamia - Hakea - Grevillea - Finschia
- Stamme Macadamieae
- Subtribe Macadamiinae
- Macadamia - Lasjia - Nothorites - Panopsis - Brabejum
- Subtribe Malagasiinae
- Subtribe Virotiinae
- Subtribe Gevuininae
- Cardwellia - Sleumerodendron - Euplassa - Gevuina - Bleasdalea - Hicksbeachia - Kermadecia - Turrillia
- Subtribe Macadamiinae
Referanser
- Weston, PH (2007). "Proteaceae". Kubitzki, K. (redaktør). Familiene og slektene til vaskulære planter. IX. Blomstrende planter - Eudicots . Springer-Verlag: Berlín. ISBN 978-3-540-32214-6.
- Hoot, SB & Douglas, AW (1998). "Phylogeny of the Proteaceae based on atp B and atp B- rbc L intergenic spacer region sekvenser". Australsk systematisk botanikk . 11 (4): 301–320. doi : 10.1071/sb98027 .
- Ramsey, HP. (1963). "Kromosomtall i proteaceae". Australian Journal of Botany . 11 (1): 1–20. doi : 10.1071/BT9630001 .
- Watson, L. & Dallwitz, MJ (1992). "Familiene til blomstrende planter: beskrivelser, illustrasjoner, identifikasjon og henting av informasjon. Versjon: 29. juli 2006" . Hentet 31. januar 2007 .
- Brown, R.Om Proteaceae of Jussieu Proceedings of the Linnean Society 10: 15-216.
Eksterne linker
-
 Data relatert til Proteaceae på Wikispecies
Data relatert til Proteaceae på Wikispecies -
 Medier relatert til Proteaceae på Wikimedia Commons
Medier relatert til Proteaceae på Wikimedia Commons - Bilder av Proteaceae fra Australian National Botanical Gardens
- Kart
{kind=link}